Engineered Extracellular Vesicles: Advancing Cancer Therapy Through Precision Nanomedicine
1School of Information Science and Engineering, Shenyang Ligong University, Shenyang 110159, China
2St. John’s Killmarnock School, Waterloo Region, Breslau, Ontario N0B1M0, Canada
3School of Mathematics and Statistics, Liaoning University, Shenyang 110036, China
4Department of Oral and Maxillofacial Surgery, Taikang Bybo Dental, Beijing 100062, China
5School of Dental Medicine, East Carolina University, Greenville, North Carolina, 27834, USA
6Department of Anesthesiology, National Cancer Center/National Clinical Research Center for Cancer/Cancer Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing 100021, China
aThese authors contributed equally to this work.
*Correspondence to: Shuang Ma, School of Information Science and Engineering, Shenyang Ligong University, Shenyang 110159, China, E-mail: 1025085193@qq.com; Jiaxuan Wu, School of Information Science and Engineering, Shenyang Ligong University, Shenyang 110159, China, E-mail: jiaxuanw@sylu.edu.cn; Yibo Gao, Department of Oral and Maxillofacial Surgery, Taikang Bybo Dental, Beijing 100062, China/School of Dental Medicine, East Carolina University, Greenville, North Carolina 27834, USA, E-mail: yibgao@126.com; Tao Yan, Department of Anesthesiology, National Cancer Center/ National Clinical Research Center for Cancer/Cancer Hospital, Chinese Academy of Medical Sciences and Peking Union Medical College, Beijing 100021, China, E-mail: yantao@cicams.ac.cn
Received: November 12 2025; Revised: January 27 2026; Accepted: April 4 2026; Published Online: May 12 2026.
Cite this paper:
Ma S, Niu Y, Sui S et al. Engineered Extracellular Vesicles: Advancing Cancer Therapy Through Precision Nanomedicine. BIO Integration 2026; 7: 1–34.
DOI: 10.15212/bioi-2025-0208. Available at: https://bio-integration.org/
Download citation
© 2026 The Authors. This is an open access article distributed under the terms of the Creative Commons Attribution License (https://creativecommons.org/licenses/by/4.0/). See https://bio-integration.org/copyright-and-permissions/
Abstract
Extracellular vesicles (EVs) are nanoscale membrane structures secreted by cells that contain proteins, nucleic acids, and lipids, and reflect the physiologic state of the parent cells. EVs have a critical role in intercellular communication, signal transduction, and tumorigenesis, influencing tumor progression, metastasis, and remodeling of the tumor microenvironment. Recent advances have highlighted the potential of EVs as natural nanocarriers for cancer therapy that offer advantages, such as biocompatibility, low immunogenicity, and the ability to cross biological barriers. Engineered EVs may overcome many of the limitations of natural EVs, including the low yield, heterogeneity, and limited targeting capabilities. Engineered EVs have shown promise in preclinical studies through genetic engineering, surface modification, and optimized loading strategies in the delivery of therapeutic agents, such as CRISPR/Cas9, mRNA, siRNA, and drugs with enhanced precision and efficacy. EVs loaded with CRISPR/Cas9 plasmids targeting PARP-1 have been shown to induce apoptosis in ovarian cancer cells and increase the sensitivity to cisplatin. Engineered EVs expressing PD-1/PD-L1 blocking antibodies have demonstrated potent anti-tumor immune activity in melanoma models by reactivating exhausted T cells, highlighting the potential for use in cancer immunotherapy. These EVs have been studied in preclinical settings involving targeted therapy, immunotherapy, and combination therapies, such as chemo-photothermal approaches, with the potential to overcoming multidrug resistance and improving treatment outcomes. Despite the promise of EVs, challenges remain in large-scale production, purification, and standardization. Corollary studies are warranted to optimize EV engineering, enhance safety, and evaluate the potential for clinical translation in oncology.
Keywords
Cancer nanomedicine, engineered EVs, extracellular vesicles, targeted drug delivery, tumor microenvironment..
Introduction
Cancer is a major global public health concern. The high incidence and mortality rates, along with profound physical and psychological impact on patients, make cancer a critical area of medical research [1]. The number of new cancer cases is projected to rise substantially according to recent global estimates, reaching 35 million by 2050, which is a 77% increase from the 20 million recorded in 2022 [2]. Although conventional treatment modalities, such as surgery, radiotherapy, and chemotherapy, can partially suppress cancer progression, conventional treatment modalities are often accompanied by severe adverse effects. However, the effectiveness of conventional treatment modalities is significantly compromised by tumor heterogeneity, drug resistance, and the complexity of the tumor microenvironment (TME) [3]. Recent advances in our understanding of tumor biology, particularly the mechanisms underlying intercellular communication and the role of extracellular vesicles (EVs) as novel nanocarriers, have shown significant potential for cancer treatment [4].
EVs are nanoscale membrane structures secreted by cells that are widely present in various body fluids. EVs have a crucial role in cell-to-cell communication by transporting bioactive molecules, such as proteins, nucleic acids, and lipids, thereby affecting the functions of recipient cells [5]. EVs are ideal drug delivery vehicles owing to biocompatibility, low immunogenicity, and the ability to penetrate biological barriers [6]. EVs offer higher stability and bioavailability compared to traditional nanomaterials and can achieve precise targeting through surface modifications [7]. These characteristics give EVs unique advantages in cancer treatment, particularly in overcoming multidrug resistance (MDR), enhancing immunotherapeutic efficacy, and improving drug delivery efficiency. With advances in research, EVs have evolved from natural biological delivery vehicles to engineered EVs, demonstrating broader application prospects in targeted drug delivery, TME modulation, and multimodal cancer therapy [8]. Researchers have discovered that engineered EVs can significantly enhance the loading capacity, targeting precision, and therapeutic efficacy [9]. For example, encapsulating chemotherapeutic drugs, gene-editing tools, or immunomodulatory molecules within EVs allows for targeted delivery to tumor cells, while minimizing damage to healthy tissue [10]. Moreover, engineered EVs can modulate the TME by targeting tumor-associated macrophages, modulating immune checkpoints, or improving tumor angiogenesis, thereby enhancing immunotherapy outcomes [11]. Preclinical research has shown that engineered EVs can successfully suppress tumor growth and metastasis, yielding promising therapeutic outcomes in various cancer models [12]. These findings highlight the considerable potential of EVs as platforms for nanomedicine delivery and provide a theoretical foundation for the development of novel cancer treatment strategies [13].
In summary, engineered EVs represent a promising nanotherapeutic platform with broad application prospects in cancer therapy based on current preclinical evidence. This study explores the biological characteristics of EVs, the diverse applications in cancer treatment as demonstrated in preclinical models, existing challenges, and future directions for advancing EV-based therapies in clinical practice.
Overview of EVs
Definition, classification, and functional properties of EVs
EVs are a diverse group of nanoscale membrane structures that are naturally synthesized and actively secreted by viable human cells. EVs are found in various bodily fluids, including blood, urine, and ascites [14]. These vesicles serve as carriers for a wide array of active biological compounds, including proteins, nucleic lipids, and acids [5]. EVs enable the transfer of molecules between donor and recipient cells via various mechanisms, including receptor-ligand interactions, endocytosis or phagocytosis, and direct membrane fusion [15]. EVs are classified into distinct subgroups based on the presumed biological origin, density, and dimensions. The primary categories of EVs include exosomes, microvesicles, apoptotic bodies (ApoVs), and more recently described subtypes, such as exomeres and migrasomes [16]. Exosomes (typically 30–150 nm in diameter) are formed intraluminally within late endosomal compartments (multivesicular bodies) and are subsequently released upon fusion of multivesicular bodies with the plasma membrane [17]. Among the various subtypes of EVs, exosomes possess a number of unique properties that make them highly suitable for engineering applications. Exosomes have a well-defined biogenesis pathway and can be genetically programmed by parent cells. The small and uniform size of exosomes facilitates large-scale production and quality control. Exosomes exhibit high stability during circulation and enable efficient in vivo delivery. Based on these advantages, exosomes have become the primary platform for research involving engineered EVs. Microvesicles (typically 100–500 nm in diameter) are generated by the direct outward budding and fission of the plasma membrane. In contrast, ApoVs (50–5000 nm in diameter) are released as blebs from the plasma membrane of cells undergoing programmed cell death [18, 19]. Exomeres are a class of non-vesicular extracellular particles (< 50 nm in diameter) that lack a lipid bilayer membrane, while migrasomes are released during cell migration [18]. Migrasomes are characterized by a pomegranate-like structure with diameters ranging from 500–3000 nm and contain multiple microvesicles with diameters ranging from 50–100 nm that can encapsulate diverse biomolecules, including proteins, lipids, and RNA, to mediate intercellular communication [19]. The above classifications are primarily established on the basis of putative biogenetic mechanisms and size ranges. Nevertheless, in practical experimental settings, EV preparations isolated via approaches, such as ultracentrifugation and size-exclusion chromatography, frequently represent heterogeneous mixtures of distinct subtypes, which exhibit substantial overlap in size and density. In practical applications, clarifying the cellular origin, isolation methodologies, and enrichment strategies of EVs is more pivotal for ensuring experimental reproducibility and comparability than rigidly differentiating the biogenetic pathways.
EVs are a highly heterogeneous population. This heterogeneity is evident at multiple levels. For example, the particle sizes within a single subcategory exhibit a continuous distribution rather than uniformity. Different EVs originating from the same parent cell may carry significantly different proteins, nucleic acids (e.g., mRNA and miRNA), and lipids. This mechanism of ‘molecular sorting’ is precisely regulated by the cellular state and microenvironment. EVs are pivotal mediators of intercellular communication and play crucial roles in diverse physiologic and pathologic processes, including immune regulation and cancer progression [20]. The functional diversity of EVs stems from inherent heterogeneity, which is determined primarily by the cellular origin and pathophysiologic context in which EVs are formed [21, 22]. Although all EVs carry a common class of biomolecules, such as signaling proteins, cholesterol, non-coding RNAs, and lipids, even EVs derived from the same parental cell exhibit significant variation in specific molecular cargo. These features give rise to distinct functional subpopulations that mediate opposing biological outcomes. For example, a subset of tumor-derived EVs in cancer can promote angiogenesis and metastasis by delivering pro-angiogenic factors [23, 24]. Another subset suppresses antitumor immunity by expressing immune checkpoint molecules, such as PD-L1 [25]. This multifunctionality allows EVs to contribute to the development of various diseases, including cancer [26]. Therefore, it is of paramount importance to view EVs as functionally heterogeneous nanocarriers to understand the multifaceted roles in the treatment of diseases. Various techniques have been used to characterize individual EVs and elucidate the physical and chemical heterogeneity, including optical and mechanical nanoparticle analysis techniques, such as cryo-electron microscopy, nanoparticle tracking analysis, atomic force microscopy, and Raman spectroscopy [27]. Single-EV characterization facilitates the identification of highly efficient EV subpopulations that carry specific therapeutic molecules or targeted ligands. Single-EV characterization also serves as the gold standard for quality control and potency determination in engineered EV production. This characterization advances EV progression towards standardized, precise clinical applications.
Biological traits of EVs
The biogenesis and release of EVs involve complex molecular mechanisms. Two primary pathways are recognized for EVs biogenesis (the endoplasmic sorting complex required for transport [ESCRT]-dependent and -independent pathways), as depicted in Figure 1 [15]. ESCRT-dependent pathways regulate EV formation by facilitating the generation of intraluminal vesicles (ILVs) within multivesicular bodies (MVBs) [28]. This process is initiated with ESCRT-0 and ESCRT-I gathering ubiquitinated cargo from MVBs, followed by the recruitment of ESCRT-III subunits through ESCRT-II, culminating in EV budding [29]. The latter pathway promotes EV biogenesis via ceramide-induced negative membrane curvature or activation of G protein-coupled sphingosine-1-phosphate receptors. This mechanism is critical for transporting EVs to ILVs [30]. In addition, tetraspanin family proteins, including cluster of differentiation 81 (CD81), cluster of differentiation 63 (CD63), cluster of differentiation 9 (CD9), tumor susceptibility gene 101 (TSG101), and apoptosis-linked gene 2-interacting protein X (ALIX), participate in endosome sorting and accumulate as distinct markers on EV surfaces. Various protein families are involved in regulating EV secretion. Notably, Ras-related proteins (RAB27, RAB35, and RAB11) enhance EV secretion, whereas SNARE proteins (SNAP25 and SNAP23) facilitate membrane fusion and cellular navigation [31].
Figure 1 Schematic representation of the biogenesis pathways and subtype characteristics of EVs. Exosomes are formed within MVBs via ESCRT-dependent or ESCRT-independent pathways and are ultimately released by fusion with the plasma membrane. Microvesicles bud outwards from the plasma membrane. ApoVs are formed during apoptosis when the cell membrane breaks down and encapsulates intracellular content, such as organelles and DNA. Exomeres lack a lipid bilayer and contain high levels of glycolytic enzymes, HSP90, and small RNAs. Migratory bodies are generated by the rupture of retraction fibers during cell migration.
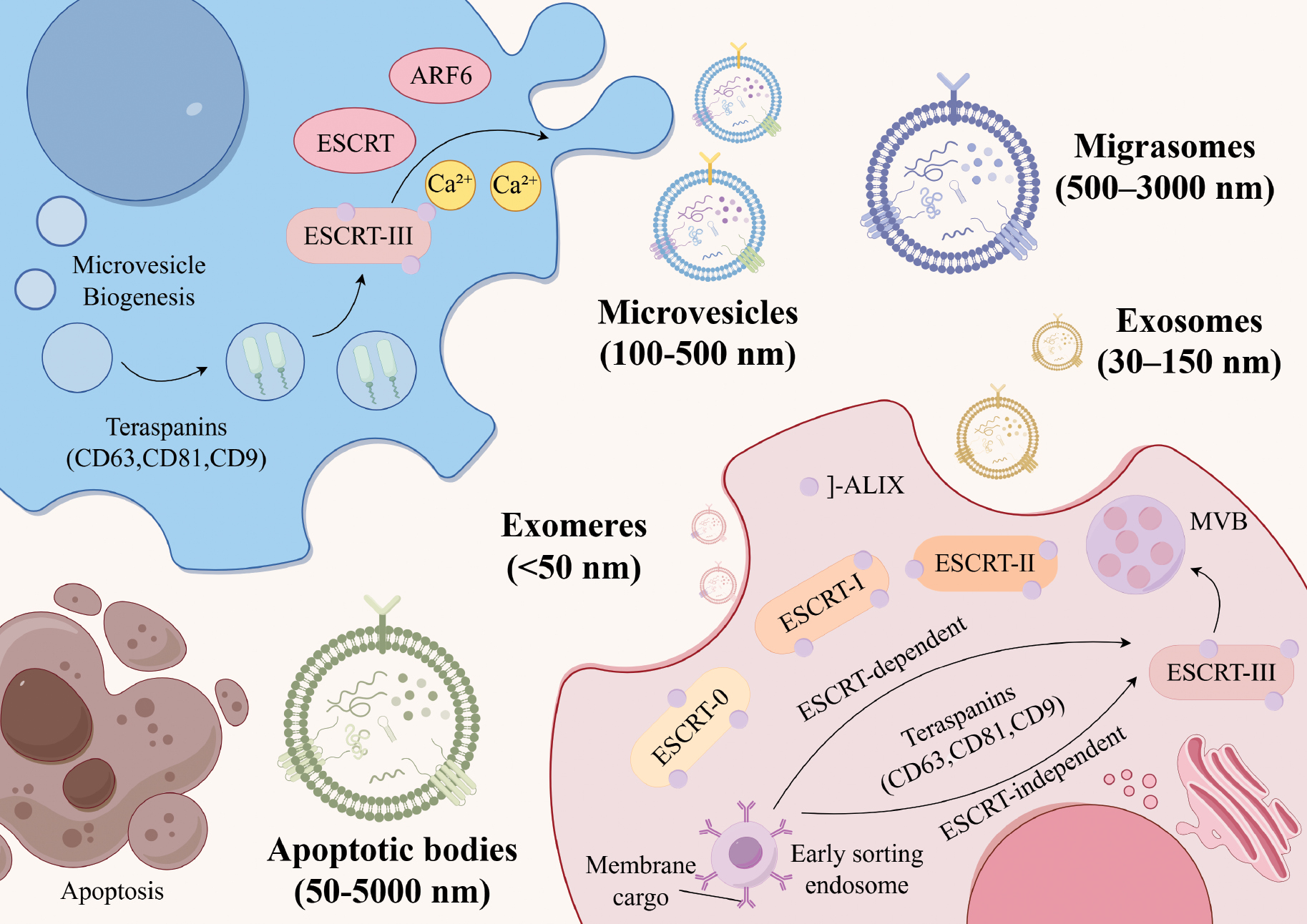
EVs exhibit a range of distinctive biological properties compared to conventional nanomaterials, including minimal toxicity, natural origin, superior biocompatibility, stability, and reduced immunogenicity. These attributes endow EVs with numerous advantages as therapeutic nanomaterials, making EVs exemplary drug delivery vehicles [32]. The natural cellular origin of EVs contributes to low immunogenicity, minimizes the risk of immune rejection, and demonstrates remarkable stability, both in vivo and in vitro.
EVs can alter their biological functions [33]. The small size and unique pattern of membrane-bound protein expression enable EVs to navigate physiologic barriers and escape clearance by the host immune system. Moreover, EVs exert considerable influence in diverse areas, including immunity, disease progression, oncogenesis, and tissue rejuvenation, primarily through intercellular vesicular transport [34]. The surfaces of EVs inherit receptors from parent cells, endowing EVs with targeting capabilities that facilitate selective interactions with specific cell types and tissue-specific drug delivery [35]. EVs demonstrate exceptional material exchangeability and tissue compatibility as drug delivery carriers, enabling precise localization and reducing drug-related toxicities. These characteristics are crucial for modulating diverse physiologic and pathologic processes, particularly intercellular communication [36]. The ability to increase the therapeutic agent concentration at target sites following systemic administration is advantageous because increasing the therapeutic agent concentration diminishes the required drug dosage for EV loading and mitigates potential toxic side effects. EV administration can bypass the P-glycoprotein (P-gp)-mediated drug efflux system, thereby attenuating drug resistance in tumor cells. Notably, EVs exhibit remarkable plasticity in terms of engineering and surface modifications [37]. Functionalization of EV surfaces is a promising approach to enhance the targeted delivery of therapeutic cargo, particularly in oncology research. Furthermore, EV modifications may yield positive outcomes in imaging research and cancer detection [38].
Role of EVs in tumorigenesis and development
EVs have a pivotal role in the occurrence, development, and metastasis of tumors. EVs participate in cell-cell communication and signal transmission and directly affect the growth and metastasis of tumor cells and remodeling of the TME. Recent studies have indicated that the role of EVs in tumor biology extends far beyond simple material transport. EVs regulate the behavior of tumor cells and dynamic changes in the surrounding microenvironment through complex mechanisms, thereby affecting tumor progression and treatment responses. The specific mechanisms underlying EVs in cell-cell communication and signal transmission, tumor growth and metastasis, and TME regulation are discussed below (Figure 2).
Figure 2 Mechanisms of EVs in tumorigenesis, development, and metastasis.
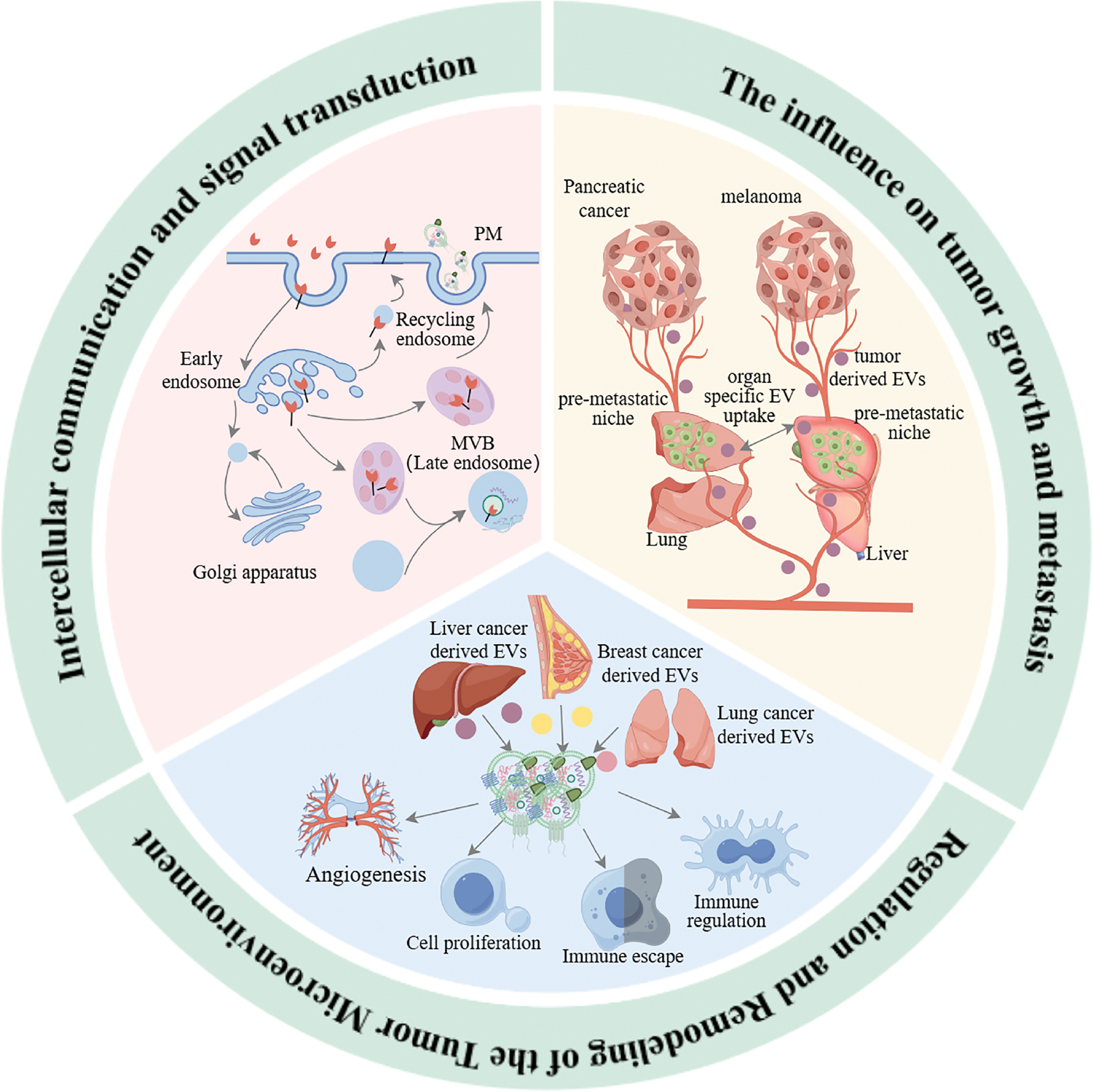
Intercellular communication and signal transmission
EVs, which are composed of phospholipids, are ubiquitously released by cells and serve as critical mediators of intercellular communication and regulation [39]. These vesicles encapsulate a carefully selected array of biomolecules, including proteins, miRNAs, mRNAs, and lipids, which confer specific functions to EVs. The structural foundation of EVs is derived from bilayer lipid molecules in the cell membranes. EVs are repositories of glycoconjugates, proteins, nucleic acids, lipids, and metabolites, and are characterized by a cellular-like topologic structure [40]. Cells actively sequester biomolecules into vesicles during apoptosis, enabling the incorporation of nucleic acids and other compounds into ApoVs. Living cell membranes generate microvesicles through direct budding from the plasma membrane, which transports bioactive molecules capable of modulating target cell behavior. The biogenesis of EVs commences with the fusion of clathrin-coated vesicles and early endosomes, progressing to the formation of multivesicular endosomes, which subsequently merge with the cell membrane to release EVs [41].
EVs are essential mediators of intercellular communication and regulation. Although non-coding RNAs (ncRNAs) constitute a vital component of EV cargo, the proportion of ncRNAs in EVs is relatively modest, particularly in mesenchymal stem cell (MSC)-derived EVs, which harbor ncRNAs associated with diverse molecular mechanisms in liver disease [42]. Virtually all mammalian cells have the capacity to secrete and uptake vesicles. As mentioned earlier, the three primary forms of EVs have different roles in tumorigenesis. ApoVs, the terminal degradation products of apoptotic cells, assist in cell-to-cell communication by transferring various materials, such as proteins, nucleic acids, organelles, and lipids, from the original cells. Studies have indicated that the ApoVs released by cells contain miR-221 and miR-222, which promote lung epithelial cell growth [43]. Using RNA sequencing technology, investigators have demonstrated that the transcriptome of ApoVs in osteoblasts at various stages closely resembles the corresponding parent cells [44]. Moreover, ApoVs released by dying stem cells contain Wnt Family Member8a protein, which stimulates nearby stem cells to internalize these vesicles and activate Wnt signaling, thereby promoting stem cell proliferation during epithelial tissue homeostasis. In addition, researchers have delineated a specific protein profile of MSC ApoVs, revealing the inheritance of apoptotic features from parental cells, such as the abundant Fas cell surface death receptor (Fas), which enhances platelet activity via Fas/FasL interactions [45]. ApoVs serve as carriers of diverse parental cell components and have important roles in various physiologic processes. Many immune cells, including dendritic cells (DCs), participate in immune responses via EV secretion [46]. EVs derived from DCs exhibit transmembrane proteins characteristic of the parental cells, including major histocompatibility complex class I, H-2-like complexes, intercellular adhesion molecule-1, membrane-activating factors, and T cell co-stimulatory factors (CD80, CD83, and CD86). These molecules stimulate immune responses in CD4+ and CD8+ T cells [47]. Likewise, EVs produced by metastatic melanoma cells that express high amounts of PD-L1 can impair CD8+ T cell function and support cancer growth through interactions with PD-1 [47].
Three primary mechanisms govern EV-target cell interactions: (i) Membrane proteins present on EVs interact with specific receptors on the target cell surface, initiating a series of intracellular signaling cascades [48]. (ii) EVs undergo membrane fusion with target cells, directly transferring the contents to the cytoplasm of the recipient cells [49]. (iii) Target cells internalize EVs through processes, such as phagocytosis [50]. These mechanisms facilitate the movement of signaling molecules and genetic material from EVs to recipient cells.
Tumor growth and metastasis
Recent studies have highlighted the crucial role of EVs in tumorigenesis initiation and progression. These microscopic structures act as channels for the transport of proteins, genetic material, and other bioactive molecules between normal and malignant cells, thereby affecting angiogenesis, metastasis, tumor initiation, growth, and immune evasion [50]. Research has indicated that EVs originating from tumors can regulate cell behavior by transmitting oncogenic proteins and genetic information [51]. For example, EVs from breast cancer cells contain miRNAs capable of activating intracellular signaling pathways, promoting the proliferation of non-cancerous cells, and the subsequent transformation into tumor cells [52]. Additionally, tumor-derived EVs from tumors have been shown to contain pro-angiogenic factors and tumor-associated antigens. These factors can directly stimulate tumor cell proliferation. When internalized by endothelial cells, angiogenesis may be activated through various mechanisms, including the hypoxia-inducible factor, Wnt, Notch, and VEGF/VEGF receptor pathways [53]. Moreover, EVs contribute to the regulation of angiogenesis-related signaling pathways by delivering specific miRNAs, thereby promoting tumor angiogenesis and facilitating the supply of oxygen and nutrients to tumors [54].
EVs have a pivotal role in the formation of pre-metastatic niches and the metastatic progression of tumor cells by facilitating the spread of oncogenic signals and promoting angiogenesis. Tumor-secreted factors recruit immune cells that enhance vascular permeability and organ tropism, thereby contributing to organ affinity [54]. Tumor-derived EVs are enriched in various biomolecules associated with cancer pre-metastatic niche formation and metastasis, including met proto-oncogene, receptor tyrosine kinase, CD97, CD151, tetraspanin, macrophage migration inhibitory factor, cell migration-inducing and hyaluronan-binding protein, C-x-C chemokine receptor (CXCR), epidermal growth factor receptor (EGFR), integrin, microRNA-105, microRNA-21, microRNA-181c, and matrix metalloproteinase-1 mRNA [55].
While aging represents a state of proliferative arrest that inhibits tumor development, mounting evidence indicates that the EV-driven aging-associated secretory phenotype exhibits complex functionality. Researchers have revealed an increased release of EVs from irradiation-induced aging of prostate cancer cells [56]. An increase in EV secretion during cellular senescence and enhanced cell growth via reverse signaling through the EphA2/ephrin-A1 pathway have also been demonstrated [57]. Notably, apoptotic EVs released by glioblastoma cells contain RNA-binding splicing factor motif 11. This factor splices cyclin D1 and MDM4, leading to the overexpression of oncogenic protein variants in target cells, which ultimately inhibits apoptosis [58]. Further studies have involved EVs derived from bladder cancer cells that can inhibit apoptosis by downregulating pro-apoptotic proteins, such as caspase 3 and Bax, and upregulating anti-apoptotic proteins, including cyclin D1 and B-cell lymphoma-2 (Bcl-2) [59]. EVs enhance the resistance of cancer cells to apoptosis by carrying specific factors or regulating the expression of key intracellular proteins.
Regulation and remodeling of the TME
The TME has emerged as a critical focus in cancer therapy. This intricate system is comprised of immune cells, tumor cells, the extracellular matrix, vasculature, and various signaling molecules. The dynamic interplay between these elements profoundly influences tumor progression, invasiveness, and treatment outcomes. Recently, EVs have garnered significant attention owing to roles in tumor development and therapeutic responses. As key mediators of information transfer within the TME, EVs significantly affect the interactions between tumor cells and the surrounding milieu, thereby affecting neoplastic growth [60]. EVs from stromal cells in the TME can affect tumor pathology by transferring the contents to nearby cells, triggering signaling events that inhibit cell death and encourage tumor growth. As depicted in Figure 3, tumor-derived EVs contribute to multiple facets of cancer progression by modifying neighboring cells, stimulating angiogenesis, conferring chemoresistance, affecting cancer-associated muscle wasting, modulating organogenesis, facilitating immune escape, orchestrating pre-metastatic niche development, promoting metastasis, and altering adipose tissue [61]. The pro-tumorigenic effects of cancer-derived EVs can also be mediated by mesenchymal cells. Bebelman MP et al. demonstrated that EVs secreted by prostate cancer cells induce myofibroblast differentiation, promote angiogenesis, and accelerate tumor growth in vivo. These effects are mediated by membrane-associated TGF-β on vesicle surfaces [52].
Figure 3 Regulation and remodeling mechanisms of EVs in the TME. (A) EVs in the neurogenic niche affect neurogenesis and neural maturation with NSCs regulating astrocyte and angiotensinogen activities by secreting miRNAs. (B) Tumor cell-derived EVs in the immune microenvironment promote immune escape, angiogenesis, and tumor progression, whereas microglia-derived EVs affect neurogenesis and neural maturation. (C) Role of EVs in the perivascular niche, where endothelial cell-derived EVs and tumor-associated macrophages affect angiogenesis and the TME by secreting factors, such as VEGF.
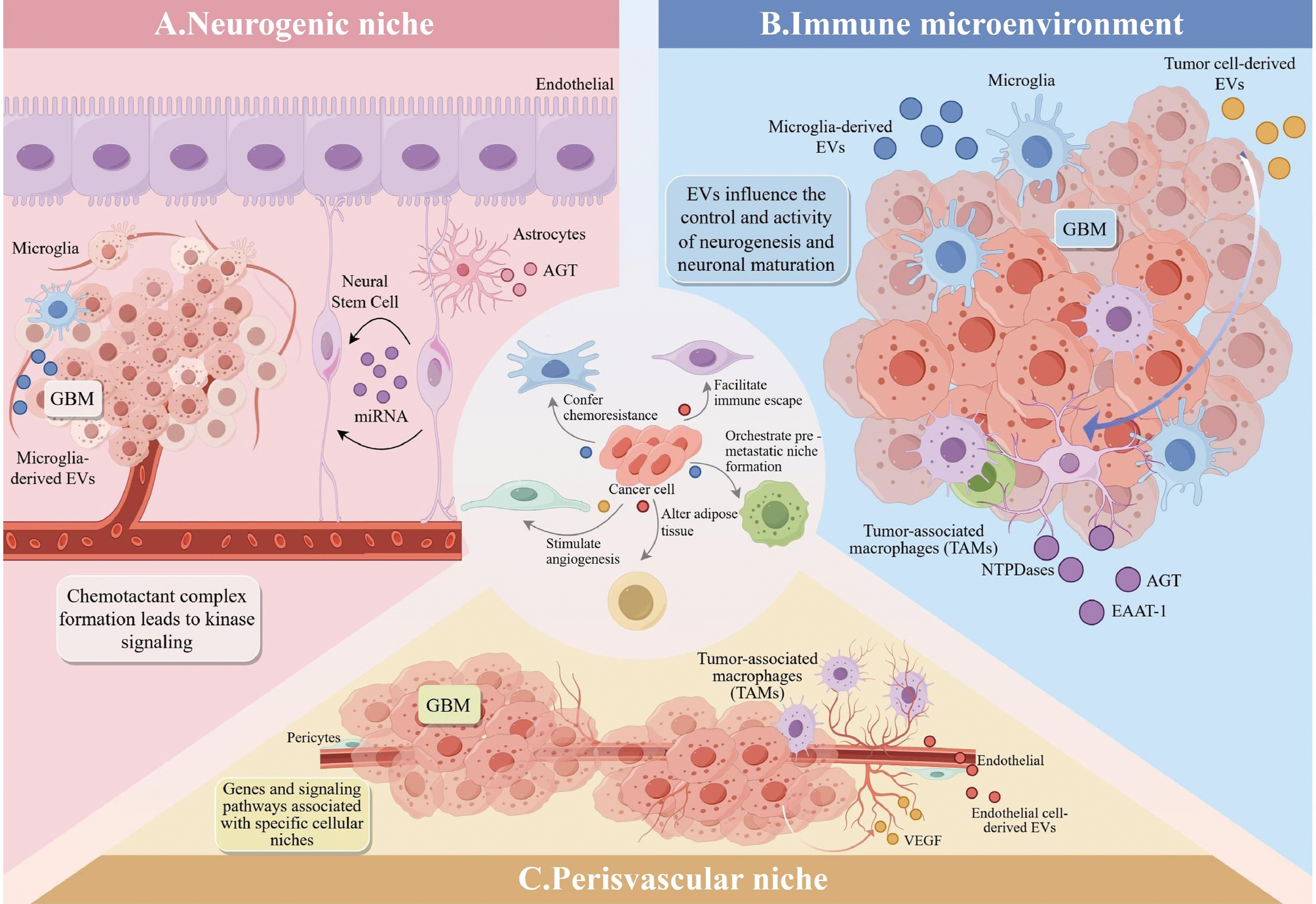
EVs from glial stem cells (GSCs) release miRNAs that exhibit selective affinities for microglia, suggesting a potential immunologic relevance. Specific RNA molecules within GSC-derived EVs, such as miRlet-7, miR-26a, miR-9, and miR-181c, have been identified and linked to microglial morphology and function [62]. Research has indicated that EVs from neural stem cells (NSCs) in the subventricular zone (SVZ) can trigger immune and inflammatory response-related transcriptional programs, functioning as microglial morphogenetic agents [63]. These NSC-derived EVs serve as communication channels between the neurogenic niche and the immune microenvironment. Studies have demonstrated that EVs can augment the CD11b+ microglial population and stimulate cytokine secretion in the SVZ. Microglia and astrocytes are crucial components of the immune microenvironment in intracranial tumors. The plasticity during neurogenesis allows microglia and astrocytes to modulate the neurogenic niches. Cytokines released by microglia significantly influence adult neurogenesis by promoting various brain inflammatory processes [64]. Reactive astrocytes contribute to the bidirectional regulation of neurogenesis. Astrocyte-derived EVs contain enzymes, such as NTPases and EAAT-1, that influence NSC proliferation and GSC differentiation. These EVs also transport neuroglobin, a neuroprotective protein indicated in brain injury models, to enhance GSC proliferation via the Wnt signaling pathway [65]. Notably, transformed astrocytes can transfer O6-alkylguanine DNA transaminases to neighboring glioma cells via EVs. This transfer is clinically significant because of the association with temozolomide resistance in glioblastoma multiforme (GBM) treatment [66]. EVs regulate tumor progression, immune escape, and TME remodeling via various mechanisms. Although these actions can promote tumor development, the actions can also be harnessed by immune cells to inhibit growth. These mechanisms of action provide new targets and strategies for tumor therapy [67].
Non-stem cell-derived EVs also influence the cellular mechanisms within the neurogenic niche, particularly in GBM. GBM cells recruit and modify non-tumor cells to regulate the TME [68]. Studies have indicated that NSCs can migrate towards gliomas and subsequently disseminate throughout the tumor bed, promoting tumor progression [69]. EVs have pivotal roles in intercellular communication through diverse mechanisms. The specific effects of EVs depend on the cellular microenvironment, which determines the cargo content. Experiments have demonstrated that NSC cultures exposed to glioblastoma-derived EVs exhibit migration rates and proliferation patterns like the transformed cells [70]. This observation provides evidence that GBM cells can transmit biological information to local NSCs, potentially inducing carcinogenic transformation. The frequent occurrence of recurrent GBM in white matter border regions, such as the SVZ, and the poorer prognosis observed in patients with GBM in which the tumors contact the SVZ (compared to tumors without SVZ contact) suggest that EVs may have a role in elucidating and potentially contributing to this phenomenon [71].
Benefits and potential of engineered EVs for cancer treatment
Although EVs have a complex role in tumor development, the inherent properties, such as low immunogenicity, biocompatibility, and ability to traverse barriers, offer unique advantages for therapeutic applications. However, native EVs also have limitations, including low yield, high heterogeneity, and insufficient targeting capacity. To address these issues, researchers have begun engineering EVs to enhance the therapeutic potential in cancer treatment. Consequently, engineered EVs have emerged in recent years, significantly improving targeting precision, drug loading efficiency, and therapeutic efficacy through genetic engineering, surface modification, and drug delivery strategies.
Pros and cons of engineered EVs
EVs serve as critical mediators of intercellular communication and have demonstrated substantial potential in both fundamental research and clinical translation. EVs can be classified into natural and engineered EVs based on whether the origins involve deliberate human design and modification. Natural EVs are nanoscale membrane vesicles that are spontaneously formed and released by various cell types (including eukaryotic and prokaryotic cells) through intrinsic biogenesis pathways under physiologic or pathologic conditions. The production of natural EVs is entirely dependent on endogenous cellular molecular mechanisms without any intervention by external genetic engineering or chemical modification [9]. Engineered EVs refer to customized vesicles generated through active biotechnological interventions involving the directed modification of EVs or the parental cells to confer specific functionalities or enhance intrinsic properties. The central objective of these engineering strategies is to overcome the functional limitations of natural EVs and achieve precise control over targeting capability, therapeutic efficacy, or diagnostic potential [72].
The core advantages of natural EVs are exceptional biocompatibility and functional integrity. Natural EVs are derived entirely from endogenous cellular components and exhibit minimal immunogenicity and high safety profiles in vivo. These vesicles harbor comprehensive molecular information reflective of parental cells under specific physiologic or pathologic states, which confers inherent potential for multi-target, multi-pathway synergistic modulation and is particularly suited for complex systemic processes, such as immune microenvironment homeostasis. The preparation workflow emphasizes efficient, non-destructive isolation and purification protocols, offering relative procedural simplicity and cost-effectiveness. However, natural EVs have significant limitations. The intrinsic compositional and functional heterogeneity results in substantial batch-to-batch variations, posing formidable obstacles to standardized scalable production. The absence of active targeting mechanisms restricts the distribution to passive accumulation, resulting in inadequate target-site enrichment and undesirable off-target accumulation. Furthermore, the uncontrollable nature of the therapeutic cargo composition and dosage precludes the precise dose titration required for potent, indication-specific therapies. In particular, industrial scalability of mammalian cell-derived EVs is severely constrained by inherently low manufacturing yields and prohibitively expensive culture conditions [6].
Engineered EVs are distinguished by the exceptional capacity for functional customization and broad adaptability to diverse clinical scenarios. Precise active targeting capabilities can be conferred through rational design strategies, significantly enhancing drug enrichment and therapeutic efficacy at the lesion sites. These vesicles enable the efficient encapsulation and delivery of diverse exogenous therapeutic payloads, including chemotherapeutic agents, nucleic acids, and proteins, facilitating synergistic or sequential therapeutic regimens for intensified intervention against complex diseases. Surface engineering offers the potential to enhance the ability to traverse stringent physiologic barriers, thereby expanding the therapeutic prospects for central nervous system disorders. In addition, the utilization of genetically tractable and scalable microbial systems for EV production offers a viable route for cost-effective large-scale manufacturing. However, advances in engineered EVs entail considerable technical complexities and inherent safety concerns. The intricate manufacturing processes prolong the development timelines and escalate costs. Genetic editing may introduce unpredictable genetic hazards, whereas chemical modifications can alter the intrinsic physicochemical properties of EVs, potentially eliciting immunogenic responses or compromising the native functions. Furthermore, the more complex origins of heterogeneity impose stringent challenges on quality control, batch-to-batch consistency, and long-term safety assessment, consequently complicating regulatory approval pathways [12].
The birth of engineered EVs
Comprehensive investigations of the potential applications of natural EVs in cancer therapy have identified numerous limitations that significantly hinder clinical application and industrial advancement. These challenges have necessitated the exploration of novel strategies to address the deficiencies of natural EVs, while capitalizing on the therapeutic benefits in cancer [73]. Engineered EVs have also been developed. These engineered EVs are optimized and regulated through human intervention and modification to address the limitations of natural EVs and endow natural EVs with enhanced characteristics tailored to meet specific application requirements. Engineered EVs offer significant advantages and potential across various domains compared to the natural counterparts.
The core advantage of engineered EVs is functional programmability, which is primarily achieved through genetic modification of cell sources and surface modification to enhance targeting ability. Active and passive loading of diverse cargos has also been performed. Table 1 summarizes the differences among various methods in terms of yield, targeting precision, load capacity, and biosafety [10]. The two major technical pathways on which genetically engineered cell sources primarily rely are lentiviral vector-mediated gene transduction and CRISPR/Cas9 system-mediated gene editing [74]. The lentiviral pathway enables the efficient loading of target molecules, such as miRNAs or membrane fusion/targeting proteins, and endogenous cellular expression through stable integration of exogenous genes. The technical core of this process lies in the design of gene constructs that carry specific signal sequences driven by strong promoters. Ultimately, the cellular biosynthetic mechanisms enrich the modified molecules within the secreted EVs. The efficiency of modification in this pathway is primarily influenced by the ExoMotif sequence design, promoter strength, strategies for selecting stable cell lines, and EV secretion capacity of the parental cells [75]. The CRISPR/Cas9 pathway involves precise genome editing, such as the removal of disease-causing genes and insertion of specific functional sequences. The key to efficiency lies in constructing an efficient ribonucleoprotein complex that can be encapsulated in EVs. This process is achieved by modifying the Cas9 protein with lipids, such as palmitoylation, and combining the Cas9 protein with targeted sgRNA. The efficiency of this pathway in editing and delivery is constrained by several factors. First, chromatin accessibility at the sgRNA target site is important with open regions being more amenable to genome editing. Second, the stoichiometric ratio of Cas9-to-sgRNA within EVs is important because an imbalance limits the formation of functional complexes in target cells. In addition, the loading strategy for EVs, the cellular DNA repair mechanisms, and the yield of EVs are important [76].
Table 1 Comparative Engineering Technologies for Engineered EVs
| Technical Category | Genetic Engineering | Surface Finish | Passive Loading | |||||
|---|---|---|---|---|---|---|---|---|
| Gene Transduction | Gene Editing | Chemical Coupling | Membrane Fusion | Electroporation | Ultrasonic Processing | Incubation Method | Refrigerating Cycle | |
| Specific methods | Lentiviral vectors carrying the ExoMotif-mir-133 sequence can stably transduce vascular smooth muscle cells, enabling sustained secretion of EVs with a high miRNA-133 payload. | The N-myristoylated Cas9 protein was co-encapsulated with sgRNA into EVs. | Targeted functionalization of EVs was achieved by sulfonating β1-adrenergic receptor antagonists (e.g., atenolol and nifedipine) to generate sulfonated derivatives. These were then covalently linked to free cysteine residues on the surface of EV membrane proteins via thioester bond formation. This strategy has two principal functions: first, it preserves the key structural domain responsible for drug binding to β1-adrenergic receptors; second, it enables the targeted modification of EVs. | The Tat-PEG-lipid (C9/C12/C14) compound was used to induce the fusion of EVs with the lipid membrane, thereby forming the EV-lipid hybrid. | The application of high-voltage electrical pulses has been demonstrated to induce poration in EV membranes, thereby facilitating drug loading. | The pharmaceutical compound or therapeutic agent should be amalgamated with the EVs, following which ultrasonic treatment should be conducted with an ultrasonic probe. | The concept of passive drug delivery is based on the dynamic remodeling of cell membranes. | EVs undergo cycles between freezing and room-temperature thawing, utilizing ice crystal formation to temporarily disrupt their membrane structure. |
| Yield | The magnitude was elevated to a degree approximately 67.7 times higher than that of the unmodified EVs. | The efficiency of gene editing can reach 25%. | Particle size and concentration stability | Higher, superior to the traditional PEG fusion method | Lower, depending on the upstream cell culture and isolation methods | The recovery rate was relatively high with approximately 98% of EVs being recovered after ultrasound treatment. | lower | The output is characterized by its stability; however, it exhibits suboptimal load efficiency. |
| Targeting accuracy | HABP actively targets calcified areas | High specificity | Significantly improves myocardial cell-specific delivery | Not evaluated, but EVs possess inherent targeting potential | – | – | – | – |
| Loading Capacity | Multiple therapeutic molecules co-loaded | The loading of the Cas9 protein was moderate. | – | Liposomes can be utilized for the encapsulation of multiple drug types, although this process has not yet been quantified. | Middle | High | Middle | Low |
| Safety Profile | Low immunogenicity, no significant toxicity | Low immunogenicity, no risk of genomic integration | No significant toxicity | Mild conditions, excellent biocompatibility of materials | It has been hypothesized that this may damage the membrane structure of EVs. | It has been hypothesized that this may affect the integrity of EV membranes. | Membrane damage was minor. | It is non-invasive and preserves the integrity of the EV membrane. |
| In vivo efficacy | A 70% reduction in calcification | It has been demonstrated that the substance under investigation has the capacity to inhibit the proliferation of prostate cancer cells that are sensitive to and resistant to enzalutamide. | Effective in ex vivo cardiac models but requires in vivo validation | No in vivo experiments were performed. | Drug-loaded EVs can effectively deliver to target cells in vivo | Loaded EVs exhibit enhanced therapeutic effects in vivo | This can enhance cellular sensitivity, but its efficacy may be lower than that of other methods. | Suitable for various types of small-molecule drugs or RNA loading |
| Stability | No significant impact on bio-stability | Not easily degradable | 80°C stable storage | Stable structure, controllable particle size, and well-defined storage conditions | Middle | Middle | Middle | High |
| Extendibility | Suitable for future large-scale production | More secure, cost-controlled, and suitable for large-scale production | Suitable for large-scale applications | Standardized operation, suitable for large-scale production | Suitable for laboratory scale | Suitable for medium-scale production | Suitable for small-scale laboratories | Easy to standardize and scale up production |
| Cost | Higher | Lower | Higher | No specific cost analysis provided | Higher | Lower | Lower | Lower |
| Ref. | [64] | [65] | [69] | [70] | [71] | [67] | [170] | [62] |
Surface modification for enhanced targeting involves the direct modification of isolated EVs to strengthen the ability to home in on pathologic sites. Mainstream approaches include chemical conjugation and membrane fusion/hydrophobic insertion methods. For example, click chemistry in chemical conjugation can generate sulfonated derivatives of β1-adrenergic receptor antagonists via sulfonation. These derivatives are then covalently attached to free cysteine residues on the surface of membrane proteins on the EV surface via thioester bond reactions, achieving targeted functionalization of the EVs. This strategy preserves the key structural domains for binding drugs to β1-adrenergic receptors [77]. Furthermore, research has demonstrated the efficient fusion of EVs and liposomes via Tat-PEG lipids, formed by coupling Tat peptides with PEG liposomes. This strategy utilizes the membrane-penetrating capacity of the Tat peptide and the hydrophobic insertion properties of lipid anchors to enable close contact and fusion of EVs and liposome membranes under mild conditions, resulting in structurally stable hybrid vesicles. This approach preserves the natural membrane composition and biological functions of EVs, while endowing EVs with the drug-loading flexibility and surface modifiability of liposomes. This offers a novel pathway for constructing multifunctional targeted delivery systems [78]. The reactivity group density of EV membrane proteins, the ligand-to-crosslinker ratio, reaction conditions, and efficacy of purification steps in removing unreacted ligands are key factors influencing ligand modification efficiency. These modifications significantly improve the ability of EVs to recognize and bind to tumor cell surface receptors, enabling targeted therapy while minimizing off-target toxicity [73].
Engineered EVs are highly efficient delivery vehicles for diverse therapeutic molecules. Passive loading involves genetically engineering parental cells to overexpress target mRNA, miRNAs, or proteins, thereby enabling natural encapsulation during EV biogenesis. Active loading involves targeting isolated EVs and using techniques, such as electroporation, ultrasonication, incubation, or freeze-thaw cycles, to load exogenous therapeutic cargo, including drugs and nucleic acids, into the EV lumen [79]. Electroporation is an efficient physical method for EV engineering that uses high-voltage electrical pulses to create temporary pores in the EV membrane. This technique significantly enhances the loading efficiency of drugs or nucleic acids (achieving encapsulation rates up to 20%), is compatible with various molecular types and offers a high loading capacity. However, the yield is low, making electroporation suitable only for small-scale laboratory preparation. Furthermore, electroporation may disrupt the structure of the EV membrane, compromising stability and innate function. Although electroporation lacks inherent targeting capabilities, electroporation can be combined with genetic engineering strategies to achieve moderate-level targeted delivery. The overall biosafety profile is favorable, although optimization of electroporation parameters is required to minimize damage to EV structure and function [80]. Ultrasonic processing is an efficient method for loading drugs into EVs. Drugs are first mixed with EVs and the mixture is then subjected to brief, low-temperature pulsing with an ultrasonic probe to disrupt membrane structures and facilitate drug entry into the vesicle lumen. The mixture is then incubated to restore membrane integrity. EV recovery rates up to 98% are possible, making this technique suitable for small molecules, proteins, and nucleic acids. While ultrasonic processing is significantly more efficient than traditional methods, such as co-incubation, power and duration require optimization to prevent membrane damage or aggregation [79]. The incubation method involves co-culturing drugs with donor cells and utilizing the natural uptake and secretion pathways to passively incorporate the drugs into EVs. This approach is the most straightforward and cost-effective, yielding EVs with intact membrane structures and low immunogenicity. However, the incubation method has a low drug-loading capacity, long processing times, and significant batch-to-batch variability. Consequently, the incubation is not suitable for scenarios requiring macromolecules or high doses but is ideal for rapid early-stage laboratory validation or when used in conjunction with other enhancement strategies [81]. The freeze-thaw cycle method involves repeated freezing and thawing at room temperature to disrupt the membrane structure of EVs, facilitating payload incorporation. This approach is simple, cost-effective, and highly biocompatible. However, the limitations include low loading efficiency, compromised membrane integrity, susceptibility to drug adsorption on the surface, and poor reproducibility of the results. While suitable for initial laboratory research, the freeze-thaw cycle methods struggles to meet the clinical demands for efficient and stable loading [82]. Notably, the conceptual framework of “engineered EVs” encompasses a diverse range of vesicle systems with distinct origins and construction strategies. To enhance scientific rigor, engineered EVs can be broadly classified into three categories. The first category comprises directly modified native EVs, which are engineered post-secretion from native cells via techniques, such as chemical conjugation or drug loading. The second category refers to parent-cell-engineered EVs that are obtained by genetically or metabolically modifying donor cells prior to vesicle biogenesis, thereby yielding vesicles with desired surface functionalities or cargo assembly. In addition to these biologically derived vesicles, the third category consists of synthetic or biomimetic EV mimetics, which are artificially constructed to recapitulate key features of native EVs, while offering greater design flexibility.
For example, researchers have developed a mimic of an apoptotic body (PARM), which has a surface coated with apoptotic body membranes and an outer layer of pH-sensitive PEG. This approach uses a bottom-up assembly to construct bionic vesicles with uniform dimensions and an intact membrane. This analog inherits the ‘eat-me’ targeting signal of natural ApoVs and can precisely load and release active components in response to the microenvironment. The analog synergistically regulates tumor-associated macrophage phenotypic polarization and phagocytic function, significantly inhibiting tumor growth and metastasis in vivo. These strategies substantially expand the functional diversity of EVs [83].
Faced with diverse technical routes, selecting the optimal combination to achieve specific therapeutic goals constitutes the core of EV research. Consequently, establishing a cross-strategic decision-making framework based on multi-dimensional considerations is imperative. Efficient loading and protection against degradation are prerequisites for nucleic acid-based cargos. Therefore, electroporation or genetically engineered source cells enable superior loading efficiency and delivery efficacy. In contrast, proteins or antibodies require preservation of the native conformation and biological activity. Genetic engineering by enabling parental cells to express fusion proteins for natural secretion or by inserting pre-assembled proteins into EV membranes via membrane fusion maximizes functional retention. Small-molecule drugs demand high loading capacity and controlled release. Approaches, such as sonication or co-incubation, offer operational simplicity but necessitate parameter optimization to balance loading efficiency and EV stability. Considering targeting modalities, active targeting relies on the specific interaction between surface-modified ligands on EVs and receptors on target cells. Chemical conjugation and genetic engineering serve as the primary strategies for active targeting, which can significantly enhance tumor-localized accumulation and mitigate off-target toxicity. Passive targeting leverages the intrinsic organotropism of EVs, which are suitable for preliminary investigations or therapeutic scenarios not requiring high specificity, albeit with relatively low efficiency. From the perspective of manufacturing processes and quality control, genetically engineered cells can be cultured at large scale in bioreactors, rendering the cells amenable to industrialization. Chemical conjugation and membrane fusion are applicable for pilot-scale production, while physical loading methods remain predominantly confined to laboratory scales. Genetic engineering and membrane fusion strategies typically exhibit superior uniformity for batch-to-batch consistency due to high operational controllability, whereas passive loading and freeze-thaw cycles are associated with greater variability and require stringent process control. With respect to functional stability, chemical conjugation and membrane fusion exert minimal impact on EV membrane integrity, thereby preserving native functionalities, whereas electroporation and sonication may induce membrane damage, necessitating parameter refinement. In terms of immunogenicity and safety, all strategies require validation of biocompatibility in in vivo models. Genetic engineering mandates assessment of exogenous gene integration risks, while chemical modification should avoid the introduction of immunogenic moieties. In summary, the selection of engineered EV strategies necessitates comprehensive consideration of multiple factors, including target cargo properties, targeting requirements, production scale, and safety profiles. Researchers can rationally select and optimally combine different technical routes tailored to specific application scenarios via the abovementioned decision-making framework, thereby efficiently developing potent, safe, and reliable engineered EVs and facilitating translation from basic research to clinical practice. In the future the integration of synthetic biology, microfluidics, artificial intelligence, and other emerging technologies is expected to enable more precise and modular engineering designs, heralding breakthroughs in cancer therapy.
These engineering strategies have greatly expanded the functional boundaries of EVs. Notably, the terms ‘EVs’ and ‘exosomes’ are often used interchangeably in engineering practice, reflecting the reality that exosomes currently constitute the primary engineering platform in cancer nanomedicine. However, other EV subtypes in addition to exosomes, particularly microvesicles and ApoVs, are gaining increasing attention due to unique functional advantages. Microvesicles have a larger diameter and offer superior cargo-loading capacity, making microvesicles especially suitable for the delivery of proteins or even organelles. ApoVs naturally display ‘eat-me’ signals, facilitating recognition and uptake by phagocytes, and thus exhibit unique value in immune regulation and cancer immunotherapy. Such functional diversity among EV subtypes underscores the importance of tailoring subtype-specific engineering strategies for distinct therapeutic objectives.
In addition, the selection of EV sources is critical and requires careful consideration in engineering design. Taking tumor-derived EVs as an example, although the natural tumor tropism and homologous targeting ability make tumor-derived EVs attractive delivery vehicles, the inherent characteristics also introduce non-negligible safety risks. Tumor-derived EVs naturally carry various pro-tumorigenic factors, including immunosuppressive molecules (e.g., PD-L1), pro-angiogenic factors (e.g., VEGF), oncogenic miRNAs, and metastasis-promoting proteins. Without appropriate engineering modification, such EVs may inadvertently promote tumor progression or exacerbate immune escape. Therefore, the application must be accompanied by careful risk-benefit assessment and targeted engineering mitigation strategies, such as knocking out or neutralizing oncogenic components via gene editing, shielding immunosuppressive ligands through surface modification, and ensuring the removal of contaminating oncogenic molecules through rigorous purification processes. In contrast, MSC-derived EVs are widely recognized for a favorable safety profile and immunomodulatory properties, while red blood cell-derived EVs are highly favored for scalable production and extremely low immunogenicity [84]. The selection of EV sources should strike a balance between targeting efficiency and safety according to the specific therapeutic context.
In conclusion, engineered EVs have effectively addressed the limitations associated with natural EVs, particularly in terms of yield, purity, heterogeneity, and targeting precision, through innovative modifications. These engineered EVs have exhibited notable advances in terms of enhanced yield, minimized heterogeneity, improved targeting capabilities, and broadened functional diversity. Consequently, these enhancements significantly augment the potential applications of EVs in cancer therapy. Further investigation into the mechanisms by which engineered EVs operate across various tumor types and integration with other therapeutic strategies is anticipated to unlock new possibilities for cancer treatment.
Application of engineered EVs for cancer treatment
Emerging applications of EVs in diagnosis and treatment
EVs are highly promising biomarkers in the field of liquid biopsy, driving transformative changes in cancer diagnosis, and treatment owing to a non-invasive nature, high information content, and dynamic monitoring capabilities. EVs are widely present in bodily fluids, such as blood, and are released by tumor cells. EVs carry a variety of biomolecules, including mutated DNA, specific proteins, and non-coding RNA, making EVs exceptionally promising markers for liquid biopsy. For example, detecting PDAC-specific EVs surface proteins, such as MUC1, EGFR, and TROP2, enables the establishment of a PDAC EV Score, which can facilitate the diagnosis of pancreatic ductal adenocarcinoma [85]. In breast cancer research, the detection of EV-miRNA-21 levels in serum revealed significantly higher concentrations in patients with metastatic breast cancer than healthy donors (P = 0.029) with particularly pronounced differences observed in the HER2-positive subgroup (P = 0.0005). EV-miRNA-21 could potentially be used as a non-invasive biomarker for diagnosing and monitoring tumor activity [86]. In addition, a novel nanomaterial (NaY) was used to efficiently enrich EVs from plasma. Through proteomic analysis, 38 proteins associated with lung squamous cell carcinoma were identified, which were shown to be simultaneously upregulated in tumor tissue and plasma EVs. Further screening of these proteins yielded five candidates (TUBB3, RPS7, RPLP1, KRT2, and VTN) for constructing a machine learning diagnostic model. This model demonstrated discriminatory efficacy in an independent validation cohort, achieving an area under the curve of 0.995, 100% sensitivity, and 96% specificity in the training set. Furthermore, two-step dimensionality reduction using LASSO and random survival forests was used to establish a prognostic risk score comprising six EV proteins (DPYD, GALK1, CDC23, UBE2L3, RHEB, and PSME1). This score significantly differentiated between high- and low-risk patients (P < 0.0001) [87]. Subsequent analysis of EV content has demonstrated high diagnostic accuracy in distinguishing patients from healthy controls.
Owing to the rapid advances in nanomedicine and molecular imaging technologies, engineered EVs have evolved from simple drug delivery vehicles into key platforms for image-guided therapy and integrated diagnosis and treatment. Researchers can achieve real-time, dynamic, visualized monitoring of drug distribution, targeted accumulation, and release processes within the body by co-loading imaging probes, such as fluorescent dyes, radionuclides, or magnetic nanoparticles, with therapeutic molecules or conjugating the imaging probes to the surface of engineered EVs. This allows for precise spatial and temporal regulation of therapeutic interventions, thereby enhancing efficacy and reducing adverse effects [88]. For example, labelling Lactobacillus fermentum-derived EVs carrying melanin nanoparticles with the fluorescent dye, indocyanine green (ICG)-NHS, enables real-time visualization of the in vivo distribution and targeting following oral administration using a small-animal near-infrared fluorescence imaging system [89]. In addition, researchers have developed a novel probe based on EVs (AS-EVs) that targets the STEAP1 protein and is loaded with a near-infrared fluorescent dye (S0456). This probe utilizes the specific targeting ability of AS-EVs towards prostate cancer cells that overexpress the surface STEAP1 protein to achieve efficient and selective enrichment at tumor sites in multiple mouse xenograft models. The fluorescent signal of the probe clearly delineates tumor boundaries, even in intramuscular tumors that wearere indistinguishable from the surrounding tissues. In surgical trials, fluorescence guidance using a probe enabled surgeons to achieve precise tumor resection and improved postoperative survival rates in mice [90].
In summary, EVs are a key medium for liquid biopsy and are transforming systems for the early diagnosis of cancer, molecular subtyping, and dynamic monitoring in unprecedented ways. Researchers have developed multiple highly sensitive and specific diagnostic and prognostic models by deciphering the molecular information EVs carry, such as nucleic acids and proteins, showcasing the significant potential for clinical application [91]. The convergence of nanotechnology and molecular imaging has transformed engineered EVs into an integrated diagnostic and therapeutic platform for various diseases. The functional modification and probe loading of EVs have enabled real-time visualization and precise regulation of drug delivery and therapeutic processes, providing innovative approaches for targeted tumor therapy and image-guided surgery [92].
Engineered EVs as nanocarriers for cancer therapy
Small-molecule drugs have achieved specific results in clinical practice as the primary tool for tumor treatment [93]. Nonetheless, these medications are susceptible to resistance and can cause significant side effects [94]. Therefore, researchers have begun investigating new drug delivery systems to address these issues. EVs serve as natural delivery vehicles with great potential to enhance drug bioavailability and therapeutic outcomes, while minimizing patient discomfort. Table 2 summarizes EV roles and mechanisms as nanocarriers in cancer therapies.
Table 2 Application and Mechanism of Action of EVs in Cancer Therapy
| Cargo Types/EVs Type | Specific Substances | EVs Source | Cancer Type | Mechanisms | Effect | Ref. | |
|---|---|---|---|---|---|---|---|
| Target therapy | DNA | CRISPR-Cas9 | HEK293 | Ovarian cancer | Suppress PARP-1 expression | It enhances chemosensitivity to cisplatin and induces apoptosis in ovarian cancer. | [95] |
| Minicircle DNA and plasmid DNA | Murine breast cancer cell line | Breast cancer | Convert the prodrug into a cytotoxic metabolite | Prevent the growth of tumors | [171] | ||
| mRNA | PTEN mRNA | DC | Glioma | Restore PTEN expression | Prevent the growth of tumors | [97] | |
| CD-UPRT-EGFP mRNA | HEK-293T | Glioma | Trigger tumor cell death by transforming the prodrug 5-FC into 5-FU | Prevent the growth of tumors | [172] | ||
| siRNA | Survivin siRNA | HEK293T | Colorectal, breast, and prostate cancers | Inhibit Survivin gene expression | Significant tumor growth regression | [100] | |
| CBSA/siS100A4 | Breast cancer cells | Triple-negative breast cancer | Inhibit S100A4 expression | Prevent the spread of cancer to the lungs | [173] | ||
| miRNA | miR-206 | Bone marrow MSCs | Osteosarcoma | Control the TRA2B gene and trigger cell death | Prevent the growth of tumors | [101] | |
| miR-126 | MDA-MB-231 breast cancer cell | NSCLC | Interrupt the PTEN signaling pathway | Inhibit cancer metastasis | [102] | ||
| miR-146b | MSC | Glioma | Inhibit EGFR gene expression | Prevent the growth of tumors | [103] | ||
| let-7a | HEK293 | Breast cancer | Reduce the expression of RAS and HMGA2 | Prevent the growth of tumors | [104] | ||
| miR-21 | 293 T cells | Glioma | Activate the PI3K/Akt signaling pathway | Prevent the growth of tumors | [174] | ||
| anti-miR-214 | HEK293T | Gastric cancer | Inhibit miR-214 expression | Reverse chemoresistance and inhibit tumor growth | [175] | ||
| Proteins | αCD3/αEGFR | M1 macrophage | Breast cancer | Recruit and activate cytotoxic T cells | Prevent the growth of tumors | [116] | |
| Survivin-T34A | YUSAC 2 | Pancreatic adenocarcinoma | Block the normal function of survivin | Prevent the growth of tumors | [176] | ||
| Gelonin | MDA-MB-231 | Breast cancer | Ensure that gelonin can be delivered specifically | Prevent the growth of tumors | [177] | ||
| Drugs | PAC | Bovine milk | Lung cancer | Inhibit cell proliferation | Prevent the growth of tumors | [106] | |
| M1 Exo-Ab | M1 macrophages | Breast cancer | Reprogram TAMs | Prevent the growth of tumors | [178] | ||
| MHC, CD86, αCD3 Ab, and αEGFR Ab | DC | B16-OVA melanoma | Activate T cells and increases killing ability | Promote the binding of T cell to cancer cells | [179] | ||
| Immunotherapy | NKEXOs | miRNA | NK cells | Neuroblastoma and breast cancer | Fuse with the tumor cell membrane or through the Fas ligand (FasL)/Fas-mediated apoptosis mechanism | It induces apoptosis in tumor cells. | [180] |
| SMART-Exos | CD3 and EGFR | Expi293F cells | Triple-negative breast cancer | Specifically, therapeutic molecules are delivered to cancer cells. | Induce cross-linking of T cells and EGFR+ cancer cells | [116] | |
| Exosomes | Antibodies against human CD3 and human HER2 | HEK293 | HER2-expressing breast cancer | Target T cell CD3 and cancer-associated HER2 | Enhance immune response | [117] | |
| MPs | CD63, CD9, TSG101 | Lung carcinoma cell lines | Lung cancer | Promote macrophage polarization and activate the Jak-STAT and MAPK pathways | Kill a variety of tumor cells | [118] | |
| sEVs | CUB domain-containing protein 1 | Irradiated tumor cells | Hepatoma and breast cancer | Activated PI3K-Akt signaling in CD8+ T cells | Prevent the growth of tumors | [119] |
EVs as nanocarriers for targeted therapy
The targeted delivery of DNA plasmid vectors to recipient cells in the context of therapeutic genome editing is highly promising. For example, a CRISPR/Cas9 expression vector delivered by hybrid EVs achieved efficient in vivo gene manipulation in MSCs. Several researchers have engineered an integrated CRISPR/Cas9 ribonucleoprotein delivery system using EVs, enabling efficient genome editing in diverse hard-to-transfect cell types, such as stem cells reprogrammed by humans to become pluripotent. Building on this approach, researchers have used EVs from cancer cells to efficiently transport CRISPR/Cas9 plasmids to target poly (ADP-ribose) polymerase-1, inducing apoptosis in ovarian cancer cells and enhancing reactivity to cisplatin [95].
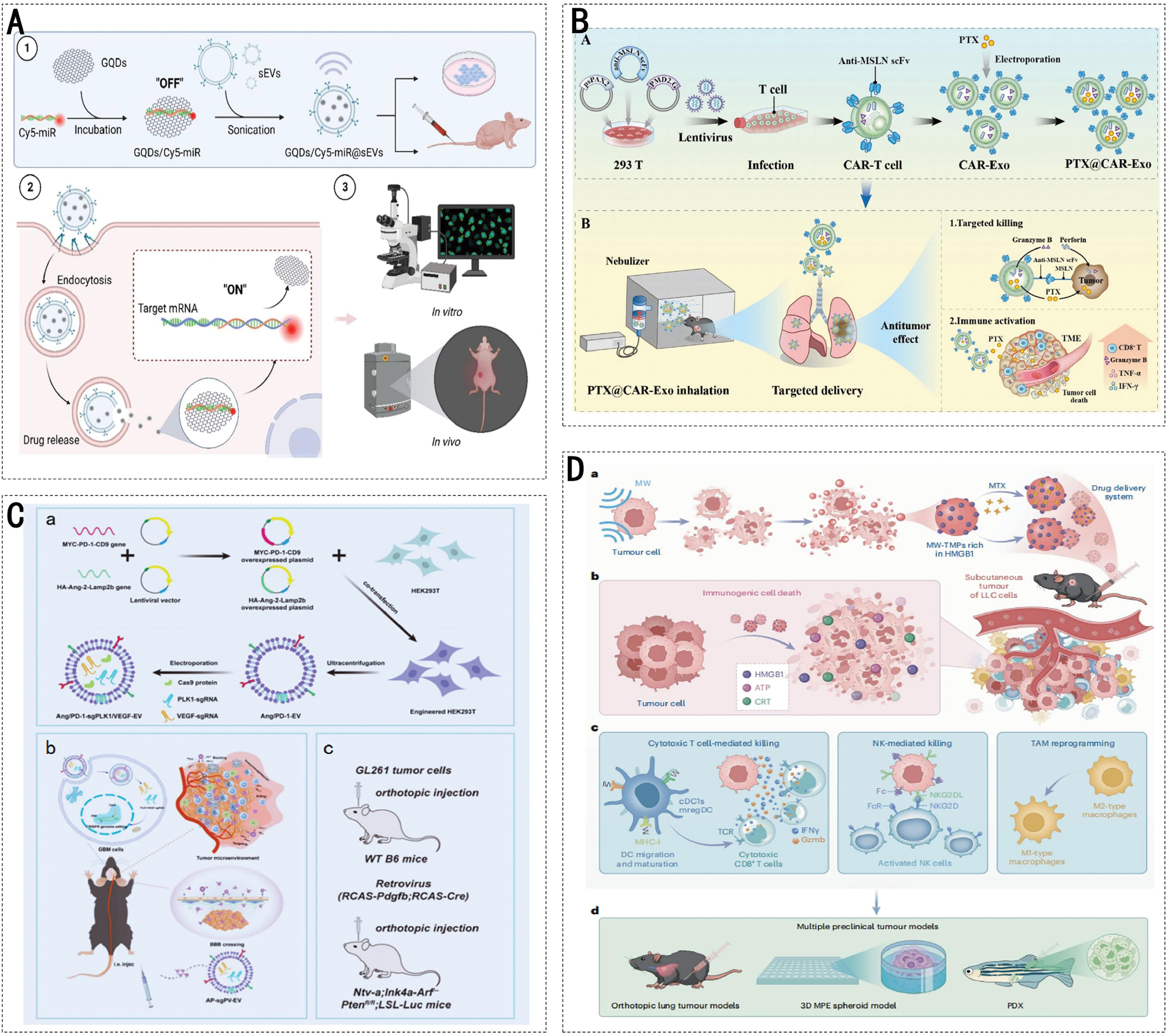
mRNA drugs for cancer treatment were actively developed following the successful development of neocoronavirus mRNA vaccines. mRNA has the potential to deliver personalized medications tailored to the unique characteristics of individual cancer cells, thereby offering hope to patients who do not respond to conventional therapies. EVs have emerged as promising delivery vehicles for therapeutic RNAs, including mRNA, siRNAs, and miRNAs [96]. Cellular nanoperforation is a novel method for producing EVs loaded with therapeutic mRNA. Several researchers have used this strategy to integrate phosphatase and angiotensin homolog mRNA into EVs, and the loading efficiency is > 1000 times higher than conventional transfection [97, 98]. Figure 4A illustrates a nanoscale fluorescent “on-off” complex composed of graphene quantum dots (GQDs) and small EVs (sEVs) engineered for targeted therapy in gastric cancer. The GQDs/Cy5-miR@sEVs complex exploits the intrinsic targeting capabilities of sEVs to deliver encapsulated miR-193a-3p specifically to gastric cancer cells, thereby enhancing precision therapy. This complex facilitates the targeted delivery of miR-193a-3p into gastric cancer cells, thereby optimizing therapeutic precision. In vitro studies demonstrated that the complex significantly downregulated CCND1 gene expression, induced apoptosis, and inhibited the proliferation and migration of gastric cancer cells. Furthermore, in vivo experiments corroborated the tumor-targeting and antitumor efficacy in a nude mouse model with no significant systemic toxicity detected. This study introduced innovative strategies and methodologies for EVs-based miRNA cancer-targeted therapy with promising clinical applications. In addition, many researchers have designed EVs to construct EXO-DEPT, which were used to deliver functional exogenous mRNA specifically targeting HER2+ cells and have been shown to block tumor growth in vivo [99]. siRNAs can knock down target gene expression in a sequence-specific manner by mediating the degradation of target mRNAs. To enhance the targeted siRNA delivery efficiency, investigators have modified EVs with folic acid and RNA aptamers as targeting ligands, enabling specific binding to receptors that are overexpressed on cancer cells. Engineered EVs modified with these ligands successfully delivered survivin siRNA to prostate and breast cancer cells, which was characterized by reduced endosomal entrapment and improved delivery efficacy [100]. miRNAs are small-molecular-weight, single-stranded ncRNAs that have important and complex roles in tumor proliferation and progression, including cell proliferation, apoptosis, tumor invasion, and epithelial-mesenchymal transition (EMT). Exosomal miRNAs can effectively inhibit tumor growth and metastasis and regulate the sensitivity of tumor cells to drugs by affecting the expression of the relevant genes. miRNAs encapsulated within EVs pass from one cell to another cell to perform biological functions. Researchers have investigated the potential of EVs as delivery systems for miRNAs in cancer treatment. HEK293T cells and MSCs are frequently used to generate EVs loaded with therapeutic miRNAs [101]. MSC-derived EVs deliver miRNAs (miR-206, 126, 146b, and let-7a) in osteosarcomas, non-small cell lung cancer (NSCLC), hepatocellular carcinoma (HCC), gliomas, and breast cancer, showing promising antitumor effects [101–104].
Figure 4 Application of engineered EVs in targeted therapy and immunotherapy. A: Targeting of tumor cells by the construction of GQDs/Cy5-miR@sEVs and application in biological systems [166]. (Figures 4A was reproduced from ref [166]. Available under a Creative Commons. Copyright 2023, The Author(s).). B: CAR-T cell-derived EVs target lung cancer cells as PTX carriers [167]. (Figures 4B was reproduced from ref [167]. Available under a Creative Commons. Copyright 2023, The Author(s).). C: AP-sgPV-EVs were constructed and applied in glioblastoma immunotherapy [168]. (Figures 4C was reproduced from ref [168]. Available under a Creative Commons. Copyright 2025, The Author(s).). D: MW-TMPs are prepared and applied to lung cancer immunotherapy [169]. (Figures 4D was reproduced from ref [169]. Available under a Creative Commons. Copyright 2025, The Author(s).).
The delivery of biologically functional proteins and therapeutic enzymes is challenging because of the cell membrane barriers. In this context, EVs have emerged as an ideal solution for protein delivery owing to a capacity to stably discharge encapsulated cargo via multiple pathways. The encapsulated cargo includes cytokines, antibodies, receptors, enzymes, antigens, and hormones. The diversity and specificity of EV-mediated protein delivery enable targeted delivery to distinct cell types, rendering the cell types applicable across various therapeutic scenarios [105].
EVs have a crucial role in drug delivery by enhancing drug stability and promoting target cell enrichment. Bovine milk-derived EVs can encapsulate and deliver PTX, and PTX-containing EVs exhibit potent antiproliferative activity against human lung cancer cells [106]. Moreover, researchers have developed PTX-containing EVs modified with aminoethylamine polyethylene glycol, which demonstrated improved anticancer efficacy and a high drug-loading capacity in a mouse model of metastatic lung cancer [107]. As shown in Figure 4B, the study involved the engineering of CAR-derived EVs expressing a second-generation chimeric antigen receptor (CAR) that targets mesothelin (MSLN). This CAR is comprised of an anti-MSLN single-chain variable fragment (scFv) and a CD8 hinge transmembrane domain. Upon inhalation-based delivery of paclitaxel (PTX)-loaded EVs, the anti-MSLN scFv on the surface specifically recognizes and binds to the MSLN antigen, which is highly expressed on the surface of lung cancer cells. This feature enables targeted delivery of drugs. Antigen-antibody binding delivers the chemotherapeutic agent precisely to the tumor site and triggers the release of granzyme B and perforin from EVs, which synergistically induces tumor cell apoptosis and remodels the tumor immune microenvironment. Studies have demonstrated that EVs can be used to deliver antibodies. For example, transfection of anti-human CD3 UCHT1 scFv antibody and the gene encoding anti-human HER2 trastuzumab scFv into Expi293F cells resulted in the production of SMART EVs. These EVs effectively and specifically triggered an immune response against tumors expressing HER2 [108]. Evidence suggests that EVs are potentially powerful tools for immunomodulation and cancer therapy. Furthermore, EVs loaded with PD-L1 blocking scFv demonstrated immunotherapeutic effects in tumor models. In addition to antibody drugs, researchers have developed a nanovaccine, exo-OVA, which uses antigens from tumors. The incorporation of neoantigens from ADPGK in M10, M16, and MC-38 tumors and B27F30 melanoma tumors into EVs was achieved using the vaccine. Among the models representing tumors, the Exo-OVA vaccine elicited strong anti-tumor immune effects [109].
When comparing delivery systems for different therapeutic molecules it is important to comprehensively evaluate delivery efficiency, stability, and clinical progress. DNA delivery systems typically have lower initial loading capacities than engineered EVs. However, owing to the natural barrier function of the EV membrane structure, the encapsulated DNA can effectively evade degradation by nucleases, resulting in significantly greater stability than free circulating DNA (e.g., NCT03228277) [110]. EVs demonstrate unique advantages in nucleic acid drug delivery. For example, EVs and EV-lipid nanoparticle (LNP) hybrid systems exhibit lower cytotoxicity and greater intestinal epithelial penetration and transport efficiency than LNPs and have favorable stability in simulated intestinal fluid. EVs and EV-LNP hybrid systems achieve 40–60% GAPDH gene silencing in Caco-2 cells, which is a significant improvement in the performance of LNPs. Furthermore, following oral administration, EVs and EV-LNP hybrid systems preferentially accumulate in the colon, demonstrating the potential for the treatment of intestinal disorders [111]. EVs exhibit enhanced biocompatibility and superior ability to traverse biological barriers in mRNA delivery. The natural membrane structure also results in prolonged circulation time in vivo, thereby effectively protecting nucleic acids. However, the core challenge facing EVs, whether for siRNA or mRNA, is the generally lower exogenous nucleic acid loading efficiency compared to artificially synthesized LNP systems. This is coupled with difficulties in large-scale production and quality control, resulting in a relatively slow clinical translation [112]. A few EV-based siRNA and mRNA projects have currently entered the early phases of clinical exploration (e.g., NCT06707961 and NCT03608631). EVs can home in on tumors and are less immunogenic, making EVs a potential vehicle for precise payload delivery. The protective function of the lipid bilayer significantly enhances miRNA stability in vivo, effectively countering enzymatic degradation (e.g., NCT07226154 and NCT07225452). In contrast, although synthetic nanocarriers allow for engineered modifications, synthetic nanocarriers often face issues related to immune clearance and inadequate targeting efficacy [113]. EVs can deliver targeted peptides or antibodies, such as anti-HER2 nanobodies, with precision, enabling active recognition and enrichment within target tissues or cells. This delivery significantly increases the drug concentration and bioavailability at the lesion site, while substantially reducing off-target toxicity to normal tissues. In contrast, delivery systems, such as liposomes and free drugs, primarily rely on passive targeting, resulting in lower efficiency and poorer specificity (e.g., NCT04288141) [114]. Small-molecule drug delivery systems, such as drug delivery systems containing PTX, doxorubicin (DOX), and tripterygin, offer the advantages of high drug loading capacity and controlled release. Drug delivery via EVs significantly increases accumulation within tumor tissues, while reducing systemic toxicity. The synergistic therapeutic potential of these systems has been validated in multiple tumor models, including lung and pancreatic cancers [115]. Overall, different therapeutic molecules have distinct advantages and disadvantages in the EV delivery system. Nucleic acid molecules (DNA, mRNA, siRNA, and miRNA) rely on the inherent protective mechanisms and targeted modifications of EVs for efficient delivery. However, loading strategies must be optimized to increase capacity. Protein-based therapeutics benefit from the natural membrane fusion capabilities of EVs, making protein-based therapeutics suitable for macromolecular delivery applications. In contrast, small-molecule drugs have a higher loading capacity and can be released in response to specific stimuli. Future efforts should focus on optimizing EVs engineering strategies that are tailored to the physicochemical properties and therapeutic requirements of different molecules, thereby advancing clinical translation.
EVs as nanocarriers for immunotherapy
In addition to targeting capabilities, engineered EVs have shown significant promise in enhancing cancer immunotherapy. AP-sgPV-EVs are genetically engineered EVs with significant potential as nanocarriers in immunotherapy, as illustrated in Figure 4C. This EV was designed to target and modulate the immune response against tumor cells by incorporating PD-1 and the brain-specific peptide angiopep-2 on the surface. In vitro and in vivo studies have demonstrated that AP-sgPV-EVs exhibit substantial antitumor effects by restoring the function of CD8+ T cells and enhancing the antitumor immune response by inhibiting the PD-1/PD-L1 signaling pathway. AP-sgPV-EVs offer a novel, highly efficient, and multi-mechanistic approach to immunotherapy and are anticipated to serve as potent therapeutic tools for the treatment of malignant tumors, such as GBM. Furthermore, researchers have proposed a mixed approach centered on natural killer (NK) cell-derived EVs assembled with a dendritic polymer core loaded with therapeutic NK cell-derived EVs with a hydrophilic shell and miRNAs (let-7a). The nanoparticle blend exhibited efficient targeting capabilities in vivo and delivered therapeutic miRNAs to these cells, leading to the simultaneous blockade of tumor growth [107]. To increase the immunogenicity of EVs, researchers from the American Chemical Society developed synthetic antibody-targeted EVs (SMART-EVs). Through the genetic presentation of two antibody varieties on EV membranes, such as monoclonal antibodies specific for CD3 on T cells and EGFR linked to cancer cells, SMART-EVs act as artificial cellular immune controllers, redirect immune effector cells, and provide potent anticancer immunity against EGFR-positive breast cancer cells [116]. The same group also generated SMART-EVs expressing anti-human CD3 and anti-human HER2 antibodies, and the resulting SMART-EVs recruited human T cells to kill HER2-positive breast cancer cells, thereby demonstrating the usefulness of this strategy in targeted cancer immunotherapy [117].
In addition to EVs from genetically engineered cells, several researchers have used EVs released by irradiated tumor cells in cancer immunotherapy. Figure 4D illustrates the generation of microwave-induced tumor-derived particles (MW-TMPs) through exposure of tumor cells to microwave radiation at a specific frequency, resulting in immunogenic cell death. MW-TMPs facilitate the activation of tumor-specific CD8+ T cells by promoting the maturation and migration of DCs, thereby enabling dual targeting of tumor cells. Furthermore, MW-TMPs reprogram tumor-associated macrophages (TAMs) within the TME, converting TAMs from the M2 phenotype, which supports tumor growth, to the M1 phenotype, which is associated with tumor suppression, thereby enhancing the antitumor immune response. The induction of immunogenic cell death by MW-TMPs in tumor cells activates the innate immune response through the release of damage-associated molecular patterns, such as high-mobility group box 1 (HMGB1), which subsequently initiates an adaptive immune response. These findings offer novel insights into immunotherapy and underscore the significant potential of MW-TMPs for tumor immunotherapy and vaccine development (Figure 4D). A recent study showed that microparticles derived from irradiated tumor cells (RT-MPs) trigger immune-stimulating death via ferroptosis. RT-MPs repolarize M2-type TAMs toward the M1-type, thereby exerting an antitumor immune response [118]. Researchers have also shown that irradiation of tumor cells leads to an increase in the immunogenicity of the EVs by boosting the manifestation of antigens associated with tumors and molecular damage patterns. These findings indicated that EVs derived from artificially treated tumor cells can provoke more robust immune responses and enhance the results of immunotherapy [119].
Engineered EVs regulate and remodel the TME
The acidic nature of the TME, potentially resulting from enhanced lactate production and glycolysis acceleration, offers a unique opportunity for the targeted modification of engineered EVs [120]. One strategy utilizes NaHCO3 (sodium bicarbonate) encapsulation within EVs, which generates CO2 bubbles upon cancer cell uptake, thereby facilitating PTX release [121]. Furthermore, a dual-responsive polymeric adhesive was designed to selectively adhere to tumor cells at pH 6.8 but remained inactive at physiologic pH (7.4) and normal body temperature (37 °C), thus playing a critical role in EV engineering [122]. Real-time monitoring of EV dynamics under various conditions uses imaging techniques, including reporter gene systems, vesicle labeling strategies, and microscopic imaging.
Recent studies have highlighted the detrimental impact of the tumor immunosuppressive microenvironment on immunotherapy efficacy, prompting an increased focus on EV-mediated TME modifications. Macrophages exhibit phenotypic plasticity in response to environmental cues, primarily manifesting as antitumor M1 and tumor-promoting M2 subtypes. M2 phenotype TAMs constitute the predominant population of immunosuppressive cells. Reprogramming macrophages from an M2-like to an M1 phenotype is a promising strategy that can be accomplished solely through the utilization of engineered EVs or in combination with other interventions [123]. For example, the signal transducer and activator of transcription 6 (STAT6) has a crucial role in M2 phenotype regulation [124]. EVs originating from M1 macrophages loaded with STAT6-targeted antisense oligonucleotides (designated as exoASO-STAT6) induce upregulation of nitric oxide synthase 2 in syngeneic models of colorectal cancer and HCC. This M1 macrophage signature aids in reshaping the TME and promotes the activation of an adaptive immune response mediated by CD8 T cells [125]. In addition to targeting TAMs, fibroblast activation protein (FAP)-α and cancer-associated fibroblasts (CAFs) are viable targets for TME modulation using engineered EVs. Tumor-derived EVs specifically targeting the FAP gene elicited targeted cytotoxic T-lymphocyte responses against tumor cells and FAP-positive CAFs, thereby reshaping the immunosuppressive TME across various models (Figure 4) [126].
Engineered EVs strategies for modulating TME are currently being validated in multiple clinical trials. These strategies exhibit diversification of targets, carriers, and delivery routes (Table 3). Currently, engineered EV therapies targeting STAT6-reprogrammed macrophages are being investigated in clinical trials. For example, Codiak BioSciences initiated a Phase I study (NCT05375604) evaluating engineered EVs derived from the HEK293 cell line and loaded with STAT6-targeting antisense oligonucleotides. These agents are administered intravenously to treat advanced HCC and other cancers. The mechanism of action involves reprogramming macrophages from the immunosuppressive M2 phenotype to the proinflammatory M1 phenotype. In addition to the STAT6-targeting strategy, a completed Phase I/II study (NCT04592484) initiated by Codiak BioSciences involved the intraluminal injection of HEK293-derived EVs loaded with a STING agonist directly into tumors. This approach aims to induce M2-to-M1 polarization and act on CD8+ T cells to treat advanced solid tumors. An ongoing Phase I/II trial at the MD Anderson Cancer Center (NCT05559177) involves the intravenous infusion of EVs derived from bone marrow-derived mesenchymal stromal cells. These EVs are loaded via electroporation with siRNA that targets KrasG12D with the aim of inhibiting downstream proliferative signaling and remodeling the TME to treat metastatic pancreatic cancer.
Table 3 Clinical Trials Investigating Engineered EVs Modulating the Tumor Microenvironment
| NCT Number | NCT05375604 | NCT04592484 | NCT05559177 | NCT05559177 | NCT01294072 | NCT01159288 | NCT01550523 |
|---|---|---|---|---|---|---|---|
| Phase | Phase 1 | Phase 1 Phase 2 | Phase 1 Phase 2 | Early Phase 1 | Not applicable | Phase 2 | Phase 1 |
| Conditions | Advanced hepatocellular carcinoma, gastric cancer. metastatic to liver colorectal cancer metastatic to liver | Advanced solid tumor | Metastatic pancreatic adenocarcinoma, pancreatic ductal adenocarcinoma | Recurrent or metastatic bladder cancer | Colon cancer | Non-small cell lung cancer | Malignant glioma of brain |
| EV source | HEK293 cell line | HEK293 cell line | MSCs of bone marrow | Tumor cell | Fruit | DC | Glioma cells |
| Engineering load or modification | Loaded with antisense oligonucleotides targeting STAT6 | Intracavitary encapsulation of synthetic cyclic dideooxynucleotides as STING agonists | Electroporation loading of siRNA targeting KrasG12D | The present study explored the hypothesis that co-incubation with DC/macrophage exosomes or membrane fusion occurs. | Physical co-incubation for curcumin loading | Enrich Dex with tumor-associated antigen-MHC-I/II complexes | – |
| Main Mechanism | Reprogramming from the immunosuppressive M2 phenotype to the pro-inflammatory M1 phenotype | Promote the polarization of M2 to M1 and activate CD8+ T cells | Inhibition of downstream MAPK/ERK proliferation signaling reduces tumor cell survival and reshapes the immune microenvironment | The carried IFN-γ, IL-12, and miR-155 promote M2→M1 polarization, reduce the number of Treg and MDSC, and enhance Th1-type immunity. | Induce macrophage polarization to M1 type and enhance CD8+ T cell infiltration | Dex activates initial/memory T cells via MHC-peptide and co-stimulatory signals, whereas mCTX pre-exhausts Treg/MDSC to relieve suppression. | Danger signals promote APC maturation, cross-present antigens, and systemically activate antitumor T cells. |
| Route of administration | Intravenous injection | Intra-tumoral injection | Intravenous injection | Subcutaneous injection | Preoperative oral medication | Subcutaneous injection | Intra-abdominal surgical implant |
| Sponsor | Codiak BioSciences | Codiak BioSciences | M.D. Anderson Cancer Center | Shanghai Pudong Hospital | University of Louisville | Gustave Roussy, Cancer Campus, Grand Paris | Sidney Kimmel Cancer Center at Thomas Jefferson University |
| Study Type | Interventional | Interventional | Interventional | Interventional | Interventional | Interventional | Interventional |
| Status | Terminated | Completed | Recruiting | Unknown status | Recruiting | Completed | Completed |
An ongoing Phase I study (NCT05559177) at Shanghai Pudong Hospital is investigating the use of subcutaneous injection of tumor cell-derived EVs. These EVs carry cytokines via co-incubation or membrane fusion with DC/macrophage EVs, thereby promoting M2-to-M1 polarization and enhancing Th1 immunity. A Phase I trial is currently recruiting participants at the University of Louisville (NCT01294072), in which fruit-derived EVs are administered orally. These EVs were physically co-incubated with curcumin to induce M1 polarization of macrophages and enhance infiltration of CD8+ T cells for the treatment of colorectal cancer. A completed Phase II study (NCT01159288) at the Gustave Roussy Cancer Center involved the subcutaneous injection of DC-derived EVs enriched with tumor-associated antigen-MHC complexes to activate T cells. This subcutaneous injection of DC-derived EVs with tumor-associated antigen-MHC complexes was combined with cyclophosphamide to deplete inhibitory cells for the treatment of non-small-cell lung cancer. In addition, a Phase I study (NCT01550523) completed at the Sidney Kimmel Cancer Center of Thomas Jefferson University involved the surgical implantation of glioma cell-derived EVs intraperitoneally. These glioma cell-derived EVs utilize danger signals to promote the maturation of antigen-presenting cells and systemically activate anti-tumor T cells for the treatment of malignant brain gliomas.
Application of engineered EVs in multimodal therapy
Chemotherapy is commonly used in traditional cancer treatment. However, malignant tumors exhibit significant heterogeneity in physiologic characteristics and responses to chemotherapeutic agents, making single-agent therapies often unsatisfactory. Consequently, combination chemotherapy strategies have been used to achieve higher efficacy. Using a combination of therapeutic techniques can optimize the effectiveness of each treatment, while minimizing limitations and adverse effects. Combination therapies associated with chemotherapy can counteract MDR and improve the efficacy of chemotherapeutic agents in cancer therapy. With the growth of EVs technology, numerous studies have explored the use of EVs to enhance the effectiveness of tumor destruction and minimize the negative side effects of chemotherapy by integrating chemotherapy with engineered EVs (Table 4).
Table 4 Engineered EVs for Chemotherapy-related Combination Therapy
| Scheme | EVs Sources | EVs types | Mechanism/Principle | Cargo | Effects | Refs. |
|---|---|---|---|---|---|---|
| Chemotherapy in conjunction with treatments aimed at overcoming drug resistance | Red blood cell | Mimic vesicles | Co-delivery of P-gp siRNA and DOX by engineered EVs | P-gp siRNA and DOX | Overcoming drug resistance and killed MDR tumors | [127] |
| HEK293 T cells | Exosomes | Engineered exosomes were loaded with miR-21i and 5-FU by mixing an appropriate amount of 5-FU with Exos using electroporation. | miR-21i and 5-FU | Reverse drug resistance and inhibit the cell proliferation | [128] | |
| 4T1 cells | Tumor cell-derived EVs | Anti-miR-21 was introduced into the producing cells, and the resulting GIONs and EVs were passed through membranes with 100 nm pores. | Anti-miR-214 and GIONs | Reduced DOX resistance led to an effective photothermal effect and showed outstanding T2 MR imaging | [134] | |
| UPCI-SCC-131R | Exosomes | The exosomal miR-155 inhibitor suppressed the stem-cell-like property | miR-155 inhibitor | Demonstrated chemosensitization toward cisplatin via upregulation of FOXO3a and downregulation of EMT markers | [135] | |
| HEK293T cells | Exosomes | Downregulating miR-214 and upregulating its potential target proteins | Anti-miR-214 | Reversed chemoresistance to cisplatin and inhibited tumor growth in vivo | [175] | |
| Bel7402 hepatocarcinoma cells | Exosomes | EVs originating from tumor cells can directly reduce P-gp expression. | PSINPs loaded with DOX | Enhancing tumor accumulation and penetration | [181] | |
| Monocyte-derived macrophages | Exosomes | Targets integrin αvβ3-overexpressing tumor cells | Dox and cholesterol-modified miRNA 159 | Enhancing the delivery of therapeutic agents specifically to cancer cells | [182] | |
| HEK293T cells | Exosomes | Silencing DARS-AS1 increased the sensitivity of TNBC cells to DOX by suppressing the transforming growth factor-β (TGF-β)/Smad3 signaling pathway-induced autophagy | DARS-AS1 siRNA and DOX | Enhancing the synergetic antitumor effects | [183] | |
| Colon cancer cells | Exosomes | Loading OXA and a CPT1A inhibitor, such as etomoxir into EVs | OXA and etomoxir | Enable OXA to have a powerful cytotoxic effect on tumor cells | [184] | |
| MSCs | Exosomes | The incorporation and release of CTX in MSCT-EXO can inhibit the activation of PI3K, Akt, and mTOR | CTX/TRAIL | Induce the apoptosis of SCC25 tumor cells and exert a good synergistic effect | [185] | |
| Chemotherapy combined with photothermal therapy | HEK-293 cells | I/R@E8-EVs | Targeted delivery of ICG and RRx-001 to the tumor site. Photothermal effect of ICG enhances drug release and therapeutic efficacy of RRx-001 | ICG and RRx-001 | Prevent the growth of tumors | [137] |
| H22 cells | Microparticles | Bi2Se3 acts as a photothermal conversion agent, converting laser energy into heat energy and generating a locally high temperature. | Bi2Se3 nanodots and DOX | A synergistic antitumor effect was achieved by integrating PTT with chemotherapy | [138] | |
| DC2.4 | EVs | The EVs encapsulate the chemotherapeutic drug DOX and serve as a template for the self-growth of gold nanoparticles, forming a popcorn-like structure | DOX | Improved cellular internalization, controlled drug release, enhanced effectiveness against tumors and minimized adverse effects | [140] | |
| 4T1 cells | Biomimetic exosomes | Reduce their drug resistance | ICG and DOX | ICG induced elevated body temperature to disassemble E-MSNs nanovehicles, resulting in successful chemo-photothermal treatment | [141] | |
| HeLa cells | ApoVs | Ti2N-DOX@ABs-based nanocarriers show advantages including stable structures, uniform sizes, and good biocompatibility | DOX | Reduce the cell survival rate steeply | [139] | |
| HeLa cells | Microvesicles | Co-delivery of PTA and chemotherapy drugs. | ICG and DOX | The combined impact of liberated ICG and DOX was able to destroy nearly all tumor cells | [186] | |
| HEK-293T cells | EVs | Effective breast cancer treatment through combination of chemo/PTT/PDT. | ICG and PTX | Efficiently suppressed tumor growth in vivo in breast tumor-bearing mice | [187, 188] | |
| Chemotherapy combined with gene therapy | SKOV3 cells | Cancer-derived exosomes | Exosomes as delivery platforms of CRISPR/Cas9 | CRISPR/Cas9-targeting PARP-1 | Reduced PARP-1 expression and increased sensitivity to cisplatin, leading to cancer cell apoptosis | [189] |
| Cal 27 cells | Microvesicles | Simultaneous delivery of therapeutic nucleic acids and chemotherapy drugs | PTX and Bcl-2 siRNA | Improved targeting and combination therapy for breast cancer. | [190] | |
| Lipid rapynanogel | Hybrid NVs coated with exosome membrane | Co-delivery of siRNA and chemotherapeutic drug | PTX and MDR1 siRNA | Prevent the growth of tumors | [144] | |
| Mouse M1 macrophages | M1mvs | Adopts the M1mv–DNA–QDs as a carrier for DOX and evade the nonspecific chemotherapy | DOX and miR21-responded hairpin DNA | Prevent the growth of tumors | [144] | |
| Blood | Exosomes | The lipid bilayer structure of exosomes allows them to co-load Dox and miR-21i with high-payloads | Dox and miR-21i | Prevent the growth of tumors | [145] | |
| HEK293 T cells | IL3 modified exosomes | Enhance the specificity and efficiency of delivering Imatinib and BCR-ABL siRNA to CML cells | Imatinib and BCR-ABL siRNA | Enhance drug sensitivity and inhibit tumor growth | [191] | |
| Red blood cells | EVs | The ASOs delivered by RBCEVs specifically target and inhibit miR-125b | miR-125b antagonized ASOs | Prevent the growth of tumors without observable cytotoxicity | [192, 193] | |
| GL261 glioblastoma cells | EVs from glioblastoma cells | The combination of gene editing and chemotherapy encourages the shift of macrophages to the M1 type and attracts CD8+ T cells | Dox and CRISPR-Cas9 system targeting CD47 IL-9 overexpression vector | Enhanced antitumor effects | [194] |
Combining chemotherapy with treatments that counteract drug resistance
MDR remains a primary driver of chemotherapy failure with biological underpinnings rooted in the complex intrinsic mechanisms that cancer cells develop or acquire. Comprehensive investigations have yielded diverse strategies for counteracting MDR, including the use of MDR inhibitors, RNA interference therapy, and novel anticancer agents designed to evade efflux. Engineered EVs offer versatile delivery platforms that enable co-delivery of anti-MDR agents and chemotherapeutic drugs, thereby suppressing resistance mechanisms and enhancing chemotherapeutic efficacy. A compelling example is the co-delivery of P-gp siRNA and chemotherapeutics via engineered EVs to combat drug-resistant tumors. Researchers have used a membrane disruption-self-assembly approach to generate engineered mimic vesicles as substitutes for natural EVs. These synthetic vesicles were loaded with P-gp siRNA and the chemotherapeutic drug DOX to implement a dual strategy of P-gp silencing and chemotherapy. Functional assays demonstrated that these engineered vectors overcame drug resistance by downregulating P-gp expression and synergistically eradicating MDR tumors through the combined mechanisms of DOX-induced growth inhibition and RNAi-mediated efflux pump suppression. This approach demonstrated the medical viability of EV-based co-delivery systems for overcoming MDR and optimizing cancer treatment outcomes [127]. Moreover, evidence suggested that altering tumor cell-derived EVs can overcome MDR and increase chemotherapy effectiveness. Figure 5A illustrates the development of a co-delivery system utilizing engineered EVs derived from tumor cells to concurrently deliver 5-fluorouracil (5-FU) and miR-21 inhibitors to drug-resistant colorectal cancer cells. These EVs demonstrated high efficiency in targeting drug-resistant cells, leading to a significant reduction in miR-21 expression and upregulation of the protein expression of the target genes (hMSH2 and PTEN), effectively reversing drug resistance. In vitro and in vivo experiments confirmed that this system markedly enhanced apoptosis, inhibited cell proliferation, and improved the efficacy of the chemotherapeutic agent, 5-FU, thereby overcoming MDR. This study introduced a novel therapeutic strategy for reversing drug resistance in colorectal cancer with promising potential for clinical applications [128]. The strategy of co-delivering functional small RNAs and anticancer drugs through EVs provides a potential approach to reverse drug resistance in tumor cells, thereby improving the efficacy of cancer therapy [129].
Figure 5 Application of engineered EVs in combination therapies A: Engineered EVs nanocarriers co-deliver 5-FU and miR-21i to enhance the effects of chemotherapy [128]. (Figures 5A was reproduced from ref [128]. Available under a Creative Commons. Copyright 2020, The Author(s).). B: EVs loaded with an miR-155 inhibitor were combined with cisplatin to treat oral cancer and overcome chemoresistance [135]. (Figures 5B was reproduced from ref [135] with permission from American Chemical Society. Copyright 2021.). C: I/R@E8-EVs use photothermal therapy with ICG and chemotherapy with RRx-001 to treat gastric cancer [137]. (Figures 5C was reproduced from ref [135] with permission from American Chemical Society. Copyright 2021.). D: ID@E-MSNs integrating DOX and ICG achieve synergistic chemo-photothermal therapy for tumor treatment [141]. (Figures 5D was reproduced from ref [141]. Available under a Creative Commons. Copyright 2020, The Author(s).). E: EVs loaded with the chemotherapeutic drug PTX and the gene therapy drug Bcl-2 siRNA were used to treat breast cancer [143]. (Figures 5E was reproduced from ref [143] with permission from American Chemical Society. Copyright 2017.). F: EVs loaded with the chemotherapeutic drug Dox and the gene therapy drug microRNA-21 inhibitor (miR-21i) for antitumor therapy [145]. (Figures 5F was reproduced from ref [145]. Available under a Creative Commons. Copyright 2020, The Author(s).).
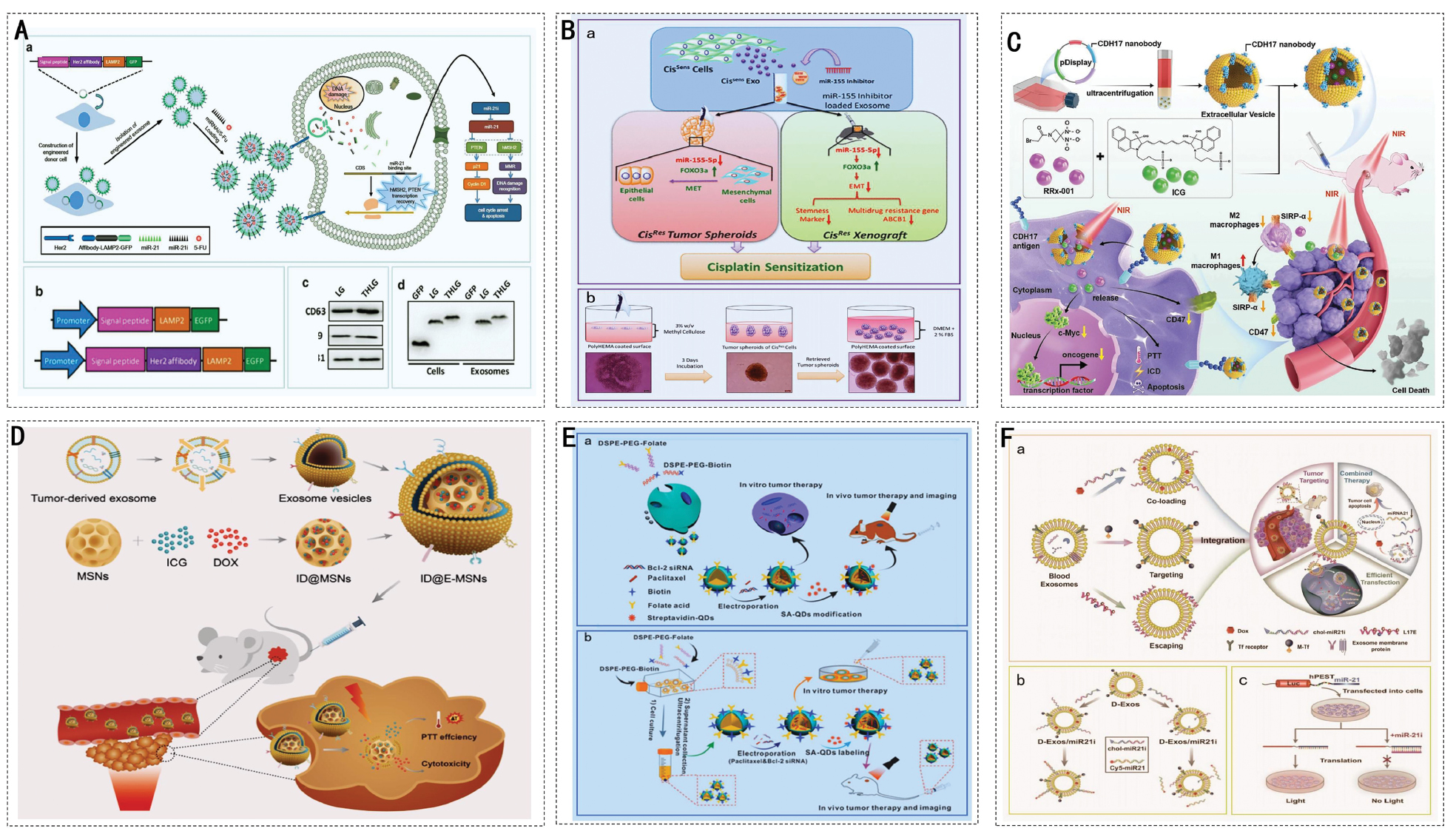
As mentioned above, dysregulated miRNA expression is a key pathologic mechanism leading to drug resistance in cancer cells. Modulating antisense inhibitors that target these miRNAs could be a potential approach for combating drug resistance [130–133]. EVs represent a promising method for RNA transport. EVs carrying anti-miR-214 effectively reversed chemoresistance to cisplatin in gastric cancer [134]. Furthermore, as shown in Figure 5B, researchers successfully reversed cisplatin resistance in oral cancer by loading miR-155 inhibitors into EVs in combination with cisplatin. This combination therapy strategy enhanced the chemotherapeutic effect by modulating the expression of FOXO3a and EMT markers and reducing the expression of stem cell properties and drug efflux transporter proteins [135]. These findings indicated that engineered EV-based delivery of anti-miRNAs and chemotherapeutic agents is a viable strategy for reversing drug resistance and enhancing chemotherapeutic efficacy.
Chemotherapy in conjunction with photothermal therapy
Photothermal therapy (PTT) is an emerging therapeutic modality in which photothermal agents (PTAs) convert light energy into heat to eliminate cancer cells. Notably, most chemotherapeutic drugs exhibit enhanced efficacy at elevated temperatures. Moderate hyperthermia can sensitize cancer cells to chemotherapy and mitigate drug resistance [136]. Consequently, the combination of chemotherapy with PTA-based PTT can synergistically enhance therapeutic outcomes, reduce chemotherapeutic dosages, and minimize adverse effects. EVs enable the co-delivery of chemotherapeutic agents and PTAs. The improved targeting and cellular internalization capabilities of engineered EVs further optimize drug utilization by reducing the chemotherapeutic dose without compromising efficacy, preventing off-target distribution of PTAs to normal tissues, and minimizing systemic toxicity of the drug. Extensive research has led to the development of various engineered EVs that significantly improve the feasibility and safety of combined chemo-PTT. Recent studies have elucidated the synergistic immune activation pathways mediated by engineered EVs. Researchers have developed EVs derived from N1-type neutrophils that overexpress interleukin (IL)-21 on the surface and are co-loaded with the photothermal agent BMT-BBT and chemotherapeutic agent oxaliplatin (OXA). The temperature-responsive release mechanism of the system was triggered by BMT-BBT in an esophageal squamous cell carcinoma model, generating heat under 808-nm laser irradiation, which rapidly released OXA upon vesicle membrane rupture. The resulting OXA-induced mitochondrial damage caused mitochondrial DNA to leak into the cytoplasm, where mitochondrial DNA was recognized by cyclic GMP-AMP synthase and catalyzed into cyclic GMP-AMP. This effect subsequently activated the STING-TBK1-IRF3 axis and upregulated IFN-β, as well as the immunogenic cell death (ICD) hallmarks of ATP efflux, CRT membrane invagination, and HMGB1 release. Concurrently, IL-21 acts on CD8+ T cells and DCs, amplifying the immune response and achieving a ‘photothermal-chemical-immune’ cascade and synergistic antitumor effect [11]. Figure 5C shows that the significant antitumor activity of EVs modified with CDH17 nanobodies (I/R@E8-EVs) comes from two processes: the ability to specifically target CDH17-positive gastric cancer cells; and photothermally triggered drug release. ICG generates a photothermal effect under near-infrared light irradiation that induces a localized temperature elevation, facilitating the controlled release of RRx-001. Furthermore, ICG-mediated PTT induces ICD by extracellularly exposing calreticulin and promoting the polarization of macrophages from the M2-to-M1 phenotype. RRx-001 inhibits the CD47-SIRPα axis to block the ‘do not eat me’ signal, thereby enhancing the phagocytosis of tumor cells by macrophages. RRx-001 also suppresses tumor proliferation by inhibiting the c-Myc pathway. These findings indicated that I/R@E8-EVs have the potential to significantly enhance the feasibility and safety of combined chemotherapy and PTT [137]. Researchers have developed systems derived from engineered cell particles for synergistic photothermal and low-dose chemotherapy. Parental cells were first modified to carry DOX and the PTT agent, Bi2Se3, then exposed to ultraviolet radiation to generate the desired particles (Bi2Se3/DOX@MPs). The findings indicated that administering Bi2Se3/DOX@MPs to mice with tumors, followed by 808-nm laser irradiation for PTT combined with low-dose chemotherapy, resulted in a significant synergistic antitumor effect [138].
The implementation of engineered EVs in a chemo-photothermal integrated treatment approach also offers the notable advantage of highly controlled drug release. Under normal physiologic conditions, chemotherapeutic agents and photothermal substances are enclosed within the membranes of specially designed EVs. Upon irradiation of the target site, PTT generates heat that rapidly increases the local temperature. This temperature increase triggers the structural disintegration of engineered EVs, enabling the controlled release of therapeutic cargo for precise cancer treatment. The in vitro photothermal chemotherapy experiments demonstrated that Ti2N-DOX@ABs, composed of Ti2N nanosheets, DOX, and apoptotic vesicles, achieved a synergistic antitumor effect. The application of Ti2N-SP@DOX alone resulted in a HeLa cell survival rate of approximately 31.3%. In contrast, the combination of Ti2N-DOX@ABs with near-infrared (NIR) laser irradiation significantly reduced the survival rate of HeLa cells to 14.4%. Live/dead cell-staining assays confirmed that Ti2N-DOX@ABs effectively eradicated HeLa cells under NIR laser exposure. Furthermore, Ti2N-DOX@ABs exhibited dual responsiveness to pH and temperature, facilitating accelerated drug release in the acidic TME and at elevated temperatures induced by the photothermal effect. This targeted drug release minimized leakage into non-target tissues and reduced side effects. Consequently, Ti2N-DOX@ABs offer an efficient, precise, and optimized synergistic therapeutic strategy for cancer treatment [139]. In addition, under NIR light irradiation, the gold nanoparticles in the engineered EVdox@AuNP structure produced a hyperthermal effect, leading to the collapse of the EV structure and prompting the rapid release of DOX. This temperature-induced structural change and drug release can be controlled and minimal amounts of drug can be released without NIR irradiation. By combining chemotherapeutic drugs and PTT, EVdox@AuNPs have demonstrated highly effective antitumor effects in in vitro and in vivo experiments, a good biosafety profile, and low side effects, resulting in precise cancer treatment [140]. As illustrated in Figure 5D, tumor cells are eradicated through the collaborative effects of PTT and chemotherapy. In addition, researchers have devised a controlled delivery system for co-loading of DOX and ICG. Tumor-derived EVs were combined with mesoporous silica nanoparticles (MSNs) preloaded with DOX and ICG, followed by the fabrication of exosome-camouflaged MSNs (ID@E-MSNs) using an extrusion technique. Similarly, the results demonstrated that ID@EMSNs could generate thermotherapy by collapsing nanocarriers, accelerating drug release, and achieving effective chemo-PTT under 808-nm NIR irradiation. In conclusion, these studies indicated that engineered EVs have significant potential as delivery platforms with strong loading capacity and good biocompatibility for combined chemo-PTT [141].
Chemotherapy paired with gene therapy
Gene therapy is a promising approach for introducing therapeutic nucleic acids into cells to amend or alter genetic information. Given that genetic mutations are a primary driver of cancer, targeting mutated genes has been shown to influence cancer treatment outcomes. Addressing these mutant genes at the genetic level through gene therapy has the potential to enhance the efficacy of cancer treatments, including chemotherapy. EVs serve as excellent nanoscale delivery systems that can simultaneously transport therapeutic nucleic acids and chemotherapeutic drugs. Furthermore, the precise delivery capability of engineered EVs can reduce chemotoxicity and unintended genetic effects in healthy cells, while also facilitating combination cancer treatments. For example, researchers co-delivered DOX and the CRISPR-Cas9 system to form EVs@Dox/sgCD47/IL-9 complexes. This combined delivery not only improved the delivery efficiency of drugs and nucleic acids but also remodeled the TME through DOX-induced DNA damage, immunogenic cell death, and gene editing. In addition, experimental results indicated that EVs@DOX/sgCD47/IL-9 exhibited significant antitumor effects in vitro and in vivo [95, 142]. The oncogene, Bcl-2, was implicated in the regulation of anti-apoptotic defense mechanisms in cells resistant to anticancer therapy, as shown in Figure 5E. Researchers have developed folate-engineered microbubbles for co-encapsulating Bcl-2 siRNA and PTX to suppress the expression of Bcl-2 and enhance chemotherapeutic efficacy. In contrast to monotherapies, this microbubble-based delivery system demonstrated superior tumor-targeting capability, effectively downregulating Bcl-2 expression, and significantly enhancing the synergistic antitumor effects of chemotherapy and gene therapy [143].
The co-delivery of functional sRNAs and anticancer drugs through EVs offers a potential pathway for reversing drug resistance. Researchers have designed biomimetic lipid/dextran-hybridized nanocarriers loaded with MDR 1-β, siRNA, and PTX. Knockdown of MDR 1 by siRNA promotes the accumulation of PTX in cells, leading to the effective inhibition of highly drug-resistant cancer cells [144]. Researchers have developed drug delivery systems that are activated upon targeting to enhance the precision of gene manipulation using EVs. EV-like vesicles derived from M1 macrophages were engineered, miR-21-responsive hairpin DNA was incorporated, and the vesicles were loaded with DOX. These modified M1 vesicles demonstrated improved specificity for drug release and potent antitumor effects [144]. To achieve precise co-delivery of drugs and nucleic acids to tumor cells, researchers have developed a nanoplatform in which DOX and cholesterol-modified miR-21i are encapsulated into blood-derived EVs, while the endosomal lysing peptide L17E and SPIONs are conjugated to the EVs membrane (Figure 5F). This design enhances endosomal escape and promotes tumor accumulation, effectively suppressing tumor proliferation in vivo. The capability of “all-in-one” nanoplatforms for cancer treatment has been demonstrated [145].
Discussion
Engineered EVs represent an emerging nanoscale drug delivery platform in the preclinical stage, the developmental trajectory of which fundamentally diverges from conventional synthetic nanocarriers. The development of traditional liposomes and polymeric nanoparticles adheres to well-defined chemical engineering paradigms, offering advantages, such as favorable batch-to-batch consistency and mature manufacturing protocols. However, these systems are constrained by inherent limitations, including suboptimal biocompatibility and elevated immunogenicity. In contrast, engineered EVs originate from natural biological systems, possess intrinsic biocompatibility and minimal immunogenicity, and have an enhanced capacity to traverse biological barriers. These characteristics confer distinctive therapeutic advantages. However, this biological provenance introduces novel challenges. The production of EVs is intrinsically a biomanufacturing process with heterogeneity significantly influenced by parental cell status, culture conditions, and isolation methodologies, thereby posing substantial difficulties in standardization and quality control. Whereas traditional nanomedicines have established relatively clear evaluation pathways at the regulatory level, engineered EVs lack globally harmonized regulatory classifications as novel therapeutic products, amplifying uncertainties in clinical translation [146].
Currently, one of the pivotal scientific challenges in the field of engineered EVs is achieving optimal synergy between targeted modification strategies and the intrinsic homing properties of EVs. EVs inherently exhibit tissue tropisms through surface-associated natural components, including integrins, immunomodulatory proteins, and glycosylation. Researchers commonly use genetic editing or chemical conjugation approaches to introduce exogenous ligands to enhance targeting specificity. However, such engineering interventions may have unintended consequences, such as excessive modification masking natural homing signals on the EV surface, altering the physicochemical properties of the membrane structure, or eliciting undesirable immunogenic responses, thereby compromising the intrinsic delivery efficiency. The risks associated with over-engineering underscore the necessity of re-evaluating the current modification paradigms. Future studies should focus on systematically elucidating the molecular mechanisms underlying EV homing from a systems biology perspective and subsequently developing minimal modification principles that enable precise delivery, while preserving the fundamental biological identity of EVs [147]. Furthermore, another fundamental controversy in this field centers on the delineation and validation of the biosafety and functional consistency of engineered EVs. Divergent cellular origins, modification methodologies, and manufacturing protocols may substantially amplify EV heterogeneity, thereby compromising therapeutic reproducibility and reliability for clinical translation [148].
In summary, although engineered EVs have demonstrated substantial potential in oncology, the transition from bench-to-bedside is impeded by multifaceted scientific, technical, and regulatory challenges. A comprehensive understanding of the fundamental distinctions from conventional nanocarriers, direct confrontation of prevailing scientific controversies, and active exploration of rational design strategies grounded in systems and synthetic biology may facilitate a paradigm shift for engineered EVs from empirical investigation toward precision engineering, ultimately establishing engineered EVs as effective modalities for next-generation precision tumor therapeutics.
Prospects and challenges of engineered EVs in cancer therapy
EVs have emerged as highly promising nanodelivery systems in preclinical research primarily because of demonstrated biocompatibility and stability in a wide range of experimental settings. Significant advances have been made in various aspects of EVs in basic and translational studies, including purification processes, biogenesis mechanisms, identification methods, and comprehensive biological characterization. Our current understanding suggests that EV-mediated cargo delivery offers protective effects against a wide array of diseases. Recent preclinical studies have increasingly highlighted the potential of appropriate modifications to significantly enhance the targeting specificity and therapeutic efficacy of EVs. Cell-free therapy based on engineered EVs has shown considerable potential and is now recognized as an innovative strategy in the field of precision medicine with highly promising applications. Notably, as research on EVs continues to advance, EVs demonstrate broad application prospects beyond tumor therapy.
Emerging trends and clinical anti-tumor
In recent years a series of breakthroughs have not only broadened the scope of EV applications in anti-tumor therapy but also accelerated the clinical translation. In addition to oncology, EV-based mRNA vaccines have demonstrated significant potential in preventing and controlling infectious diseases, particularly in the response to sudden viral outbreaks. Research has confirmed that EVs can serve as highly efficient and safe mRNA delivery platforms [149]. For example, pulmonary-derived EVs carrying mRNA that encodes the SARS-CoV-2 spike protein and is formulated as an inhalable dry powder vaccine enhances the targeted distribution and retention time of mRNA within the lungs. Pulmonary-derived EVs carrying mRNA that encodes the SARS-CoV-2 spike protein also effectively activate mucosal and systemic immune responses. This approach has been shown to achieve rapid viral clearance and provide long-lasting immune protection in mouse and monkey models [150]. EVs have been shown to contribute to the stability and delivery efficiency of mRNA vaccines. EVs protect mRNA from nuclease degradation and improve cellular uptake efficiency. EVs can also evade immune clearance via surface proteins, such as CD47. EVs can also be engineered. For example, by modifying the surfaces with targeting peptides to strengthen tissue-specific delivery capabilities, EVs provide a flexible and safe platform for developing vaccines against multiple infectious diseases [151]. Therefore, EVs exhibit great potential for the development of mRNA vaccines for non-tumor indications, especially for the design of vaccines targeting respiratory and mucosal pathogens. This offers a new technological approach for the prevention and control of future infectious diseases.
Innovations in engineering modification techniques have greatly improved the functional programmability of EVs. While traditional EVs engineering relies on gene overexpression or chemical modification, the introduction of synthetic biology tools has provided unparalleled precision and diversity in functional programming. In recent years, researchers have extensively used these tools to engineer EVs with greater precision, thereby enhancing targeting capabilities and therapeutic functions [152]. In the field of skeletal disease treatment, researchers have proposed a modular engineering strategy based on probiotic-derived EVs. Although this study used chemical modification to attach the bone-targeting peptide, SDSSD, to the surface of LGG-EVs, the overall design followed the modular assembly principle of synthetic biology. This chemical modification involves the rational integration of targeting modules (SDSSD), delivery modules (bacterial vesicles), and therapeutic modules (endogenous osteogenesis-related microRNAs) to create an engineered vesicle system capable of tissue-specific delivery and disease modulation [5]. Furthermore, researchers have employed synthetic biology strategies to construct a ClyA-BMP-2-CXCR4 fusion expression system within Escherichia coli cells, generating bacterial EVs that possess bone-targeting (CXCR4) and osteogenic induction (BMP-2) capabilities. This study demonstrated how meticulously designed genetic circuits can integrate multifunctional modules directly into the biosynthesis of EVs, enabling the efficient one-step preparation of “engineered vesicles” from “engineered cells.” This study provided a model for developing therapeutically engineered EVs with complex functions [6]. These engineered strategies, which are based on synthetic biology principles, expand the functional capabilities of EVs and provide a programmable nanoscale platform for the precise delivery of drugs and treatment of diseases.
Emerging technologies offer new solutions to the inherent challenges posed by the heterogeneity and complexity of EVs. The heterogeneity, complexity, and inter-individual variation of EVs present difficulties for EV-based therapeutic strategies [153]. The introduction of artificial intelligence (AI) and machine learning technologies has provided powerful tools for analyzing the composition of EVs, predicting functions, and optimizing treatment regimens [154]. AI can help screen EV subpopulations bearing specific biomarkers by analyzing the molecular characteristics of proteins, lipids, and nucleic acids within the EV subpopulations. AI can facilitate the development of targeted drug-delivery systems. For example, machine learning can identify characteristic proteins, such as CLTC, EZR, and TLN1, in cancer-associated EVs, enabling the construction of classification models that can accurately distinguish cancerous EVs from non-cancerous EVs. This provides a powerful tool for the early diagnosis of cancer and differentiation of cancer types. Furthermore, analyzing exosomal surface-enhanced Raman scattering spectra using deep learning models enables the simultaneous detection of multiple early-stage cancers (e.g., breast and lung cancers) and determination of the tissue origins within a single assay. This approach provides highly sensitive and specific diagnostic information for early cancer screening and has the potential to be expanded in the future to achieve more precise, personalized diagnostics. Artificial intelligence drives the precise design and functional optimization of EVs as biomarkers and drug carriers. AI also supports personalized diagnosis and treatment decisions by integrating multidimensional data, which provides a promising technological foundation for the future of personalized medicine.
Challenges and solutions
Despite the significant potential of engineered EVs, the transition from laboratory research to clinical applications is beset with numerous challenges. Currently, EV production is largely confined to the laboratory scale and relies on traditional methods, such as ultracentrifugation and polymer precipitation [155]. These approaches have low recovery rates and are time-consuming and labor-intensive. The approaches also readily introduce impurities, making it difficult to meet the stringent purity, homogeneity, and batch-to-batch consistency requirements for clinical-grade products. To advance industrialization, cGMP principles must be rigorously implemented at every stage of production. Currently, few contract manufacturing organizations can produce EVs because such production imposes specialized demands on facilities, methods, and quality control systems. Compared to traditional biological products, EV manufacturing requires additional considerations, such as viral clearance studies, EV-specific purification steps, and ensuring the physicochemical stability of the lipid bilayer structure. Companies should plan for cGMP capacity building early in the preclinical research phase and collaborate with experienced partners to design closed, single-use production processes tailored to the characteristics of EVs. This approach reduces the risk of cross-contamination and enhances the flexibility of production. A pioneering study reported the first clinical-grade engineered exosome therapeutic, termed iExosomes, established with a comprehensive GMP-compliant manufacturing process for the treatment of pancreatic ductal adenocarcinoma harboring KrasG12D mutations. To achieve scalable production, the research team implemented a series of GMP-compliant strategies. Bone marrow-derived MSCs, which have extensive clinical usage history, were selected as the exosome source with high-yield donors identified through rigorous screening. A quantum bioreactor was employed to establish a closed culture system, enabling single-run exosome yields of tens of trillions of particles. Downstream purification utilized a closed process integrating centrifugation, filtration, and ultracentrifugation, with the FDA-approved Plasma-Lyte serving as the clinical buffer. Electroporation of siRNA was upgraded to the 4D Nucleofector LV large-scale system, facilitating 20 mL-scale closed processing with direct application without wash steps. This study demonstrated that exosomes remained stable during long-term storage at −80°C, retaining biological activity and physical integrity upon thawing for defined periods. This manufacturing paradigm established quality release specifications based on NanoSight particle size analysis, flow cytometric marker detection, and in vitro potency assays, thereby providing a comprehensive GMP production framework for the clinical translation of engineered EV therapeutics [72]. Furthermore, equipment commonly used in EV production, such as three-dimensional culture systems, tangential flow filtration, and chromatographic purification, requires systematic process validation and scale-up studies to ensure stable operation within cGMP environments. This process involves setting up stable cell bank systems, optimizing cell culture processes (e.g., use of chemically defined serum-free media and three-dimensional bioreactors to increase yield and consistency), and developing efficient, scalable downstream purification technologies [156]. For example, combining magnetic stirring with tangential flow filtration can significantly increase the recovery rate of EVs and improve the purity of particles and proteins at the research scale. If this approach is integrated with chromatography steps and validated for an ability to remove host cell proteins and nucleic acid residues, the approach could offer a novel technical solution for preparing GMP-grade EVs [157]. Moreover, the potential of microfluidic technology has become increasingly apparent in recent years. By designing intricate microchannels, continuous and high-throughput separation and purification of EVs can be achieved. This approach significantly reduces processing time and enhances product homogeneity through precise parameter control, offering a highly promising technical pathway for large-scale production [158]. Another core challenge confronting the industrialization of EVs is the batch-to-batch variability. EV heterogeneity stems from various factors, including the state of donor cells, culture conditions, and isolation methods. These factors can lead to significant variations in particle size distribution, drug loading capacity, membrane protein composition, and biological potency between different batches. To minimize this variation, the upstream processes must be addressed, which involves the use of process analytical techniques to monitor critical parameters, such as pH, dissolved oxygen, and metabolite concentrations in real time, thereby ensuring a stable culture environment [159]. Quality control standards for downstream processes should be based on multidimensional physicochemical and biological parameters, moving beyond traditional assays of particle concentration and total protein content. To achieve the expected quality, nanoparticle tracking analysis, dynamic light scattering, high-performance liquid chromatography, proteomics, and functional activity assays should be incorporated. These methods allow for the comprehensive characterization of the key quality attributes of EVs. Equally critical is the establishment of efficacy assessment standards linked to clinical outcomes, such as cellular uptake efficiency, targeted delivery efficacy, and pharmacodynamic evaluation in animal models of disease. There is an urgent need for international collaboration within the industry to develop widely recognized reference materials and standardized testing protocols to establish a unified benchmark for quality control [160].
EVs remain subject to evolving regulatory frameworks worldwide, as complex biological products. Authorities have yet to agree on uniform guidelines for classification as cell or gene therapy products or novel drug delivery systems, definitions of impurities (particularly process- and product-related impurities), and safety assessments (including immunogenicity, off-target distribution, and long-term toxicity). Companies must engage with regulatory bodies, such as the US FDA or China NMPA, during the early stages of research and development (R&D) to reach a consensus on the validation of production processes, design of non-clinical studies, and quality control strategies based on risk assessment principles. Robust data demonstrating the stability, safety, and intended functionality of the modifications must be provided for engineered EVs, along with assessments of potential immunogenicity. Mitigating immunogenicity and off-target effects is crucial for the clinical safety of engineered EVs. These risks can be reduced by selecting humanized donor cells or utilizing cell-free synthetic systems [161]. Furthermore, computer simulations and AI-assisted design can optimize the binding specificity between the targeted ligands and receptors, thereby predicting and mitigating off-target risks. In terms of delivery strategies, developing stimulus-responsive release systems enables precise and controlled drug release at target sites, thereby reducing exposure to healthy tissues, such as pH, enzyme, or photothermal-sensitive formulations [162].
EVs constitute an emerging therapeutic product category for which no unified global regulatory guidelines have yet been established. From the early stages of R&D, enterprises must maintain close communication with regulatory authorities to reach a consensus on critical issues, such as manufacturing processes, quality control, and non-clinical study design. Particular attention should be paid to scientifically grounded risk assessments concerning EV classification, impurity characterization, and viral safety evaluation rather than simplistically applying existing biological product guidelines. Industry organizations should actively promote interdisciplinary and cross-departmental dialogue platforms to jointly develop technical guidance principles applicable to EVs products, thereby paving the way for their compliant development and market entry [161].
Based on a comprehensive analysis of these challenges and controversies, we propose two forward-looking research directions with substantial translational potential. First, the development of intelligent responsive modular EV engineering platforms is required. Whereas current engineering strategies are predominantly static, future efforts should focus on constructing intelligent EV systems capable of dynamically responding to pathologic microenvironmental cues. This effort necessitates the convergence of materials science, synthetic biology, and EV technology, exemplified by integrating environmentally responsive modules into EV architectures to achieve triggered drug release at specific sites, or alternatively, designing logic-gated systems inspired by synthetic biology principles that activate therapeutic functions only when multiple disease-specific criteria are simultaneously satisfied. Second, the establishment of AI and multi-omics-driven rational design frameworks for EVs. Confronted with the complex heterogeneity of EVs, traditional trial-and-error approaches exhibit limited efficiencies. A data-driven paradigm should be constructed employing machine learning to integrate multi-omics data with functional validation outcomes, thereby establishing predictive models to guide optimal EV subpopulation selection and engineering design. This should be coupled with microfluidic and single-vesicle analysis technologies to develop high-throughput screening platforms that accelerate the discovery and optimization of engineered EVs with desired performance characteristics.
Through comprehensive elucidation of the fundamental distinctions between engineered EVs and conventional nanocarriers, direct confrontation of prevailing scientific controversies, and active exploration of forward-looking research directions, this field is poised to achieve a paradigm shift from empirical investigation to rational design. Ultimately, such progress could pave the way for the successful clinical translation of engineered EVs as next-generation precision nanomedicines, thereby providing innovative breakthrough strategies for oncology pending rigorous safety and efficacy validation.
Potential failure modes and risks
Despite the enormous therapeutic potential of engineered EVs, the clinical translation is confronted with systemic risks and potential failure modes that must be rigorously addressed, and these challenges persist throughout the entire product lifecycle. The most fundamental challenge arises from inherent heterogeneity and the associated risks of batch consistency. Even for cells of the same origin under identical engineering modification and culture conditions, subtle fluctuations in epigenetic status and microenvironment can lead to remarkable differences in RNA cargo and protein composition of the secreted vesicles. Furthermore, off-target effects of gene editing, heterogeneous efficiency of chemical modifications, and limitations of current isolation techniques further amplify the variability of the final product, resulting in a highly complex mixture. Such heterogeneity directly leads to inconsistent batch-to-batch performance in terms of size distribution, drug loading capacity, and biological activity, constituting the core risk responsible for the uncertainty and irreproducibility of clinical efficacy [163, 164]. In the pursuit of precise delivery, targeting efficiency and off-target accumulation represent another critical contradiction. Excessive engineering may mask the natural “don’t-eat-me” signals on the vesicle surface, leading to rapid clearance. Meanwhile, the introduced targeting ligands may also compromise binding affinity due to spatial conformation issues. Even after modification, engineered EVs may still undergo non-specific accumulation in organs, such as the liver and spleen, owing to the nanoscale size. If these organs are sensitive to the loaded chemotherapeutic drugs or toxins, severe dose-limiting toxicity may occur. For delivery of gene-editing tools, such as CRISPR/Cas9, off-target distribution implies the potential risk of accidental gene editing in non-target cells, which could lead to tumorigenesis [79]. Furthermore, engineering modifications may fundamentally alter the low immunogenicity of EVs. Heterologous components from parental cells, media additives, non-natural moieties introduced by chemical conjugation, or loaded exogenous proteins may all serve as antigens recognized by the immune system, resulting in the production of anti-drug antibodies, allergic reactions, or even cytokine storms. Such immunogenic risks are particularly critical for therapeutic strategies requiring long-term and repeated administration. More profound and unknown risks arise from long-term toxicity. Delivered nucleic acid drugs may exert persistent and unintended interference with intracellular gene regulatory networks. In addition, the long-term metabolic fate of inorganic nanomaterials or non-degradable polymers introduced by certain engineering strategies remains unclear; these materials may accumulate in specific organs and induce chronic inflammation. These nanoscale carriers may also be internalized by normal tissue cells, causing long-term and unpredictable impacts on the tissue microenvironment and stem cell niches [165]. Therefore, the clinical translation of engineered EVs must be grounded in a profound understanding of the potential risks. Future research should shift the focus from mere functional enhancement to a comprehensive assessment balancing risks and benefits, and risk control strategies should be integrated into engineering design during the early stages of R&D.
Data availability statement
The original contributions presented in this study are included in this article. Further inquiries should be directed to the corresponding author.
Ethics statement
No direct interactions with human or animal subjects were involved. Therefore, ethical approval and informed consent were not required.
Author contributions
Shuang Ma, Yibo Bao and Tao Yan conceived the study, designed the methodology, and oversaw the entire project. Yue Niu and Wanying Xu performed the literature review, manuscript translating, and data pre-processing. Jiaxuan Wu and Xiaolin Wu contributed to theoretical framework and interpretation. All authors approved the final manuscript, with Shuang Ma and Tao Yan providing final edits and supervision.
Acknowledgements and funding
This work was supported by National Natural Science Foundation of China (Grant Nos. 2022JC2K01, 82073327, and 62541321), Doctoral Research Start up Fund of Shenyang University of Technology (Grant Nos. 1010147001258), Liaoning Provincial Natural Science Foundation Doctoral Research Launch Project (Grant No. 2025-BS-0346), the Applied Basic Fund of Liaoning Province (Grant No. 2025-BS-0346), CAMS Innovation Fund for Medical Sciences [CIFMS] (2025-I2M-C&T-B-047), National Cancer Center Climbing Fund (NCC202404005), the Bethune Charitable Foundation (BCF, kqxs-2024A-02), the National Science and Technology Major Project of China [Innovation 2030] (2024ZD0533300 and 2024ZD0533303), and the National Key Research and Development Program of China (2023YFC3403804). The authors thank Home for Researchers (www.home-for-researchers.com). The Graphical abstract, Figure 1 and Figure 3 were created by Figdraw. https://www.figdraw.com.
Conflict of interest
The authors declare that they have no conflicts of interest.
Graphical abstract
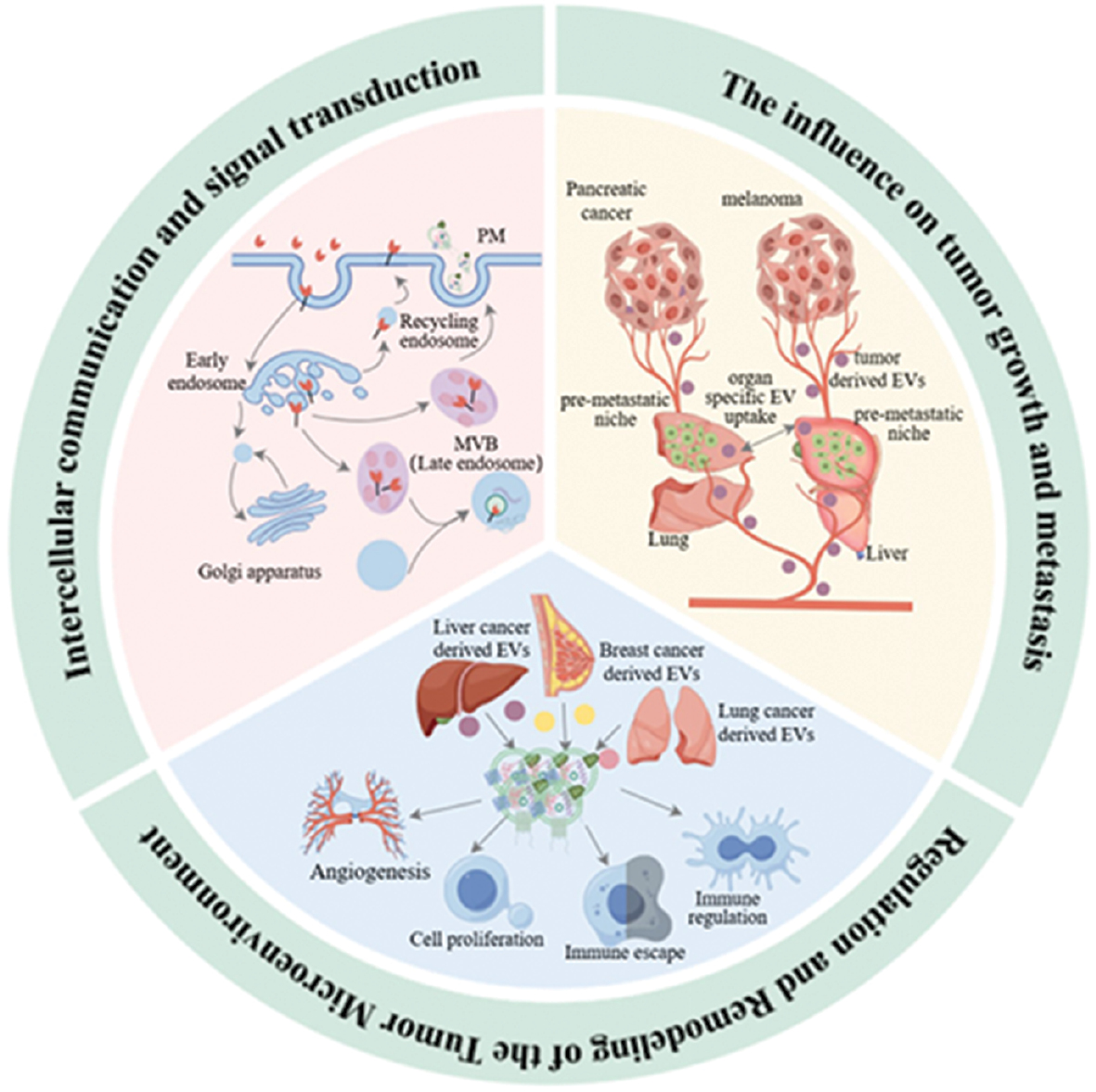
Highlights
- Extracellular vesicles serve as biocompatible nanocarriers for efficient therapeutic delivery in cancer.
- Engineering strategies enhance EV yield, targeting specificity, and cargo-loading efficiency.
- Engineered EVs enable precise gene and immune modulation with potent antitumor activity.
- Engineered EVs show strong potential for use in personalized and next-generation cancer therapies.
In brief
Engineered extracellular vesicles (EVs) have emerged as a promising platform in cancer nanomedicine based on preclinical findings, offering biocompatible and low-immunogenic delivery systems capable of crossing biological barriers. This study summarizes EV biology, engineering strategies, and therapeutic applications, highlighting advances in genetic modification, surface functionalization, and cargo loading demonstrated in in vitro and in vivo studies. Engineered EVs may enable the precise delivery of CRISPR/Cas9, mRNA, siRNA, and chemotherapeutics with potential to enhance targeted therapy, immunotherapy, and combination treatments, while overcoming drug resistance in preclinical models. Despite the challenges in scalable production and standardization, engineered EVs hold significant potential for advancing precision oncology and clinical translation.
References
- Santucci C, Carioli G, Bertuccio P, Malvezzi M, Pastorino U, et al. Progress in cancer mortality, incidence, and survival: a global overview. Eur J Cancer Prev 2020;29(5):367-81. [PMID: 32740162 DOI: 10.1097/cej.0000000000000594]
- Casolino R, Tatah L, Charnaud S, Santero M, Ilbawi A, et al. The WHO global landscape of cancer clinical trials. Nat Med 2025;31(9):2901-12. [PMID: 40926101 DOI: 10.1038/s41591-025-03926-x]
- Liu B, Zhou H, Tan L, Siu KTH, Guan XY. Exploring treatment options in cancer: tumor treatment strategies. Signal Transduct Target Ther 2024;9(1):175. [PMID: 39013849 DOI: 10.1038/s41392-024-01856-7]
- Chen Y, Du M, Yu J, Rao L, Chen X, et al. Nanobiohybrids: a synergistic integration of bacteria and nanomaterials in cancer therapy. BIOI 2020;1(1):25-36. [DOI: 10.15212/bioi-2020-0008]
- Zhou G, Zhou Q, Li R, Sheng S, Gao Q, et al. Synthetically engineered bacterial extracellular vesicles and IL-4-encapsulated hydrogels sequentially promote osteoporotic fracture repair. ACS Nano 2025;19(16):16064-83. [PMID: 40237831 DOI: 10.1021/acsnano.5c03106]
- Liu H, Song P, Zhang H, Zhou F, Ji N, et al. Synthetic biology-based bacterial extracellular vesicles displaying BMP-2 and CXCR4 to ameliorate osteoporosis. J Extracell Vesicles 2024;13(4):e12429. [PMID: 38576241 DOI: 10.1002/jev2.12429]
- Liu H, Wu Y, Wang F, Wang S, Ning J, et al. Bone-targeted engineered bacterial extracellular vesicles delivering miRNA to treat osteoporosis. Compos Part B Eng 2023;267:111047. [DOI: 10.1016/j.compositesb.2023.111047]
- Qian L, Chen P, Zhang S, Wang Z, Guo Y, et al. The uptake of extracellular vesicles: research progress in cancer drug resistance and beyond. Drug Resist Updat 2025;79:101209. [PMID: 39893749 DOI: 10.1016/j.drup.2025.101209]
- Han R, Zhou D, Ji N, Yin Z, Wang J, et al. Folic acid-modified ginger-derived extracellular vesicles for targeted treatment of rheumatoid arthritis by remodeling immune microenvironment via the PI3K-AKT pathway. J Nanobiotechnology 2025;23(1):41. [PMID: 39849554 DOI: 10.1186/s12951-025-03096-5]
- Bai M, Li Z, Shi T, Li X, Li J, et al. Exogenous extracellular vesicles as emerging platforms in translational medicine. BIO Integration 2025;6:1-25. [DOI: 10.15212/bioi-2025-0122]
- Wu W, Deng Z, Liu X, Yang Y, Yuan H, et al. Engineered N1 neutrophil-derived vesicles for photothermal-enhanced immunochemotherapy of esophageal cancer. Nano Res 2025;18(10):94907965. [DOI: 10.26599/nr.2025.94907965]
- Ji N, Wang F, Wang M, Zhang W, Liu H, et al. Engineered bacterial extracellular vesicles for central nervous system diseases. J Control Release 2023;364:46-60. [PMID: 40237831 DOI: 10.1021/acsnano.5c03106]
- Luo T, Fan Z, Zeng A, Wang A, Pan Y, et al. Biomimetic targeted co-delivery system engineered from genomic insights for precision treatment of osteosarcoma. Adv Sci (Weinh). 2025;12(2):e2410427. [PMID: 39555699 DOI: 10.1002/advs.202410427]
- Jeppesen DK, Zhang Q, Franklin JL, Coffey RJ. Extracellular vesicles and nanoparticles: emerging complexities. Trends Cell Biol 2023;33(8):667-81. [PMID: 3673775 DOI: 10.1016/j.tcb.2023.01.002]
- Yu J, Sane S, Kim JE, Yun S, Kim HJ, et al. Biogenesis and delivery of extracellular vesicles: harnessing the power of EVs for diagnostics and therapeutics. Front Mol Biosci 2023;10:1330400. [PMID: 38234582 DOI: 10.3389/fmolb.2023.1330400]
- Zhou X, Huang J, Zhang D, Qian Z, Zuo X, et al. Small extracellular vesicles: the origins, current status, future prospects, and applications. Stem Cell Res Ther 2025;16(1):184. [PMID: 40247402 DOI: 10.1186/s13287-025-04330-5]
- Guo W, Li Y, Pang W, Shen H. Exosomes: a potential therapeutic tool targeting communications between tumor cells and macrophages. Mol Ther 2020;28(9):1953-64. [PMID: 32563274 DOI: 10.1016/j.ymthe.2020.06.003]
- Ashoub MH, Salavatipour MS, Kasgari FH, Valandani HM, Khalilabadi RM. Extracellular microvesicles: biologic properties, biogenesis, and applications in leukemia. Mol Cell Biochem 2024;479(2):419-30. [PMID: 37084166 DOI: 10.1007/s11010-023-04734-y]
- Ou Q, Huang W, Wang B, Niu L, Li Z, et al. Apoptotic vesicles: therapeutic mechanisms and critical issues. J Dent Res 2024;103(11):1057-65. [PMID: 39282215 DOI: 10.1177/00220345241265676]
- Sun B, Li R, Ji N, Liu H, Wang H, et al. Brain-targeting drug delivery systems: the state of the art in treatment of glioblastoma. Mater Today Bio 2025;30:101443. [PMID: 39866779 DOI: 10.1016/j.mtbio.2025.101443]
- Fyfe J, Casari I, Manfredi M, Falasca M. Role of lipid signalling in extracellular vesicles-mediated cell-to-cell communication. Cytokine Growth Factor Rev 2023;73:20-6. [PMID: 37648617 DOI: 10.1016/j.cytogfr.2023.08.006]
- Kalluri R. The biology and function of extracellular vesicles in immune response and immunity. Immunity 2024;57(8):1752-68. [PMID: 39142276 DOI: 10.1016/j.immuni.2024.07.009]
- Wang Y, Zhao M, Liu S, Guo J, Lu Y, et al. Macrophage-derived extracellular vesicles: diverse mediators of pathology and therapeutics in multiple diseases. Cell Death Dis 2020;11(10):924. [PMID: 33116121 DOI: 10.1038/s41419-020-03127-z]
- Kopec M, Abramczyk H. The role of pro- and antiangiogenic factors in angiogenesis process by Raman spectroscopy. Spectrochim Acta A Mol Biomol Spectrosc 2022;268:120667. [PMID: 34865975 DOI: 10.1016/j.saa.2021.120667]
- Galassi C, Chan TA, Vitale I, Galluzzi L. The hallmarks of cancer immune evasion. Cancer Cell 2024;42(11):1825-63. [PMID: 39393356 DOI: 10.1016/j.ccell.2024.09.010]
- Zhou G, Li R, Sheng S, Huang J, Zhou F, et al. Organoids and organoid extracellular vesicles-based disease treatment strategies. J Nanobiotechnology 2024;22(1):679. [PMID: 39506799 DOI: 10.1186/s12951-024-02917-3]
- Park J, Kim H, Roh YH, Ko J. Advances in single extracellular vesicle characterization and multiplexed profiling. TrAC Trends Anal Chem 2026;195:118588. [DOI: 10.1016/j.trac.2025.118588]
- Ye Z, Li G, Lei J. Influencing immunity: role of extracellular vesicles in tumor immune checkpoint dynamics. Exp Mol Med 2024;56(11):2365-81. [PMID: 39528800 DOI: 10.1038/s12276-024-01340-w]
- Banjade S, Zhu L, Jorgensen JR, Suzuki SW, Emr SD. Recruitment and organization of ESCRT-0 and ubiquitinated cargo via condensation. Sci Adv 2022;8(13):eabm5149. [PMID: 35363519 DOI: 10.1126/sciadv.abm5149]
- Peche VS, Pietka TA, Jacome-Sosa M, Samovski D, Palacios H, et al. Endothelial cell CD36 regulates membrane ceramide formation, exosome fatty acid transfer and circulating fatty acid levels. Nat Commun 2023;14(1):4029. [PMID: 37419919 DOI: 10.1038/s41467-023-39752-3]
- Bae JW, Yi JK, Jeong EJ, Lee WJ, Hwang JM, et al. Ras-related proteins (Rab) play significant roles in sperm motility and capacitation status. Reprod Biol 2022;22(2):100617. [PMID: 35180567 DOI: 10.1016/j.repbio.2022.100617]
- Elsharkasy OM, Nordin JZ, Hagey DW, de Jong OG, Schiffelers RM, et al. Extracellular vesicles as drug delivery systems: why and how? Adv Drug Deliv Rev 2020;159:332-43. [PMID: 32305351 DOI: 10.1016/j.addr.2020.04.004]
- Kwok ZH, Wang C, Jin Y. Extracellular vesicle transportation and uptake by recipient cells: a critical process to regulate human diseases. Processes (Basel) 2021;9(2):273. [PMID: 34336602 DOI: 10.3390/pr9020273]
- Li S, Li W, Wu X, Zhang B, Liu L, et al. Immune cell-derived extracellular vesicles for precision therapy of inflammatory-related diseases. J Control Release 2024;368:533-47. [PMID: 38462043 DOI: 10.1016/j.jconrel.2024.03.007]
- Zheng W, He R, Liang X, Roudi S, Bost J, et al. Cell-specific targeting of extracellular vesicles through engineering the glycocalyx. J Extracell Vesicles 2022;11(12):e12290. [PMID: 36463392 DOI: 10.1002/jev2.12290]
- de Jong B, Barros ER, Hoenderop JGJ, Rigalli JP. Recent advances in extracellular vesicles as drug delivery systems and their potential in precision medicine. Pharmaceutics 2020;12(11):1006. [PMID: 33105857 DOI: 10.3390/pharmaceutics12111006]
- Cheng HY, Su GL, Wu YX, Chen G, Yu ZL, et al. Extracellular vesicles in anti-tumor drug resistance: mechanisms and therapeutic prospects. J Pharm Anal 2024;14(7):100920. [PMID: 39104866 DOI: 10.1016/j.jpha.2023.12.010]
- Sabani B, Brand M, Albert I, Inderbitzin J, Eichenseher F, et al. A novel surface functionalization platform to prime extracellular vesicles for targeted therapy and diagnostic imaging. Nanomedicine 2023;47:102607. [PMID: 36167305 DOI: 10.1016/j.nano.2022.102607]
- Sánchez GB, Bunn KE, Pua HH, Rafat M. Extracellular vesicles: mediators of intercellular communication in tissue injury and disease. Cell Commun Signal 2021;19(1):104. [PMID: 34656117 DOI: 10.1186/s12964-021-00787-y]
- Skotland T, Sagini K, Sandvig K, Llorente A. An emerging focus on lipids in extracellular vesicles. Adv Drug Deliv Rev 2020;159:308-21. [PMID: 32151658 DOI: 10.1016/j.addr.2020.03.002]
- Kang T, Atukorala I, Mathivanan S. Biogenesis of extracellular vesicles. Subcell Biochem 2021;97:19-43. [PMID: 33779912 DOI: 10.1007/978-3-030-67171-6_2]
- Nemeth K, Bayraktar R, Ferracin M, Calin GA. Non-coding RNAs in disease: from mechanisms to therapeutics. Nat Rev Genet 2024;25(3):211-32. [PMID: 37968332 DOI: 10.1038/s41576-023-00662-1]
- Li M, Liao L, Tian W. Extracellular vesicles derived from apoptotic cells: an essential link between death and regeneration. Front Cell Dev Biol 2020;8:573511. [PMID: 33134295 DOI: 10.3389/fcell.2020.573511]
- Huang K, Xu Y, Feng T, Lan H, Ling F, et al. The advancement and application of the single-cell transcriptome in biological and medical research. Biology (Basel) 2024;13(6):451. [PMID: 38927331 DOI: 10.3390/biology13060451]
- Zhang X, Tang J, Kou X, Huang W, Zhu Y, et al. Proteomic analysis of MSC-derived apoptotic vesicles identifies Fas inheritance to ameliorate haemophilia a via activating platelet functions. J Extracell Vesicles 2022;11(7):e12240. [PMID: 36856683 DOI: 10.1002/jev2.12240]
- Liu A, Peng P, Wei C, Meng F, Huang X, et al. Apoptotic vesicles derived from mesenchymal stem cells ameliorate hypersensitivity responses via inducing CD8+ T cells apoptosis with calcium overload and mitochondrial dysfunction. Adv Sci (Weinh) 2025;12(22):e2407446. [PMID: 40089865 DOI: 10.1002/advs.202407446]
- Xu F, Jiang D, Xu J, Dai H, Fan Q, et al. Engineering of dendritic cell bispecific extracellular vesicles for tumor-targeting immunotherapy. Cell Rep 2023;42(10):113138. [PMID: 37738123 DOI: 10.1016/j.celrep.2023.113138]
- Kooijmans SAA, de Jong OG, Schiffelers RM, Schiffelers RM. Exploring interactions between extracellular vesicles and cells for innovative drug delivery system design. Adv Drug Deliv Rev 2021;173:252-78. [PMID: 33798644 DOI: 10.1016/j.addr.2021.03.017]
- Karmacharya M, Kumar S, Cho YK. Tuning the extracellular vesicles membrane through fusion for biomedical applications. J Funct Biomater 2023;14(2):117. [PMID: 36826916 DOI: 10.3390/jfb14020117]
- Ginini L, Billan S, Fridman E, Gil Z. Insight into extracellular vesicle-cell communication: from cell recognition to intracellular fate. Cells 2022;11(9):1375. [PMID: 35563681 DOI: 10.3390/cells11091375]
- Wang X, Yang X, Huang C, Liu T, Zang H, et al. Tumor-derived extracellular vesicle PD-1 promotes tumor immune evasion via disruption of peripheral T cell homeostasis. Cancer Lett 2025;612:217486. [PMID: 39864541 DOI: 10.1016/j.canlet.2025.217486]
- Bebelman MP, Smit MJ, Pegtel DM, Baglio SR. Biogenesis and function of extracellular vesicles in cancer. Pharmacol Ther 2018;188:1-11. [PMID: 29476772 DOI: 10.1016/j.pharmthera.2018.02.013]
- Lugano R, Ramachandran M, Dimberg A. Tumor angiogenesis: causes, consequences, challenges and opportunities. Cell Mol Life Sci 2020;77(9):1745-70. [PMID: 31690961 DOI: 10.1007/s00018-019-03351-7]
- Zhang S, Yang J, Shen L. Extracellular vesicle-mediated regulation of tumor angiogenesis—implications for anti-angiogenesis therapy. J Cell Mol Med 2021;25(6):2776-85. [PMID: 33586248 DOI: 10.1111/jcmm.16359]
- Bao Q, Huang Q, Chen Y, Wang Q, Sang R, et al. Tumor-derived extracellular vesicles regulate cancer progression in the tumor microenvironment. Front Mol Biosci 2021;8:796385. [PMID: 35059436 DOI: 10.3389/fmolb.2021.796385]
- Lehmann BD, Paine MS, Brooks AM, McCubrey JA, Renegar RH, et al. Senescence-associated exosome release from human prostate cancer cells. Cancer Res 2008;68(19):7864-71. [PMID: 18829542 DOI: 10.1158/0008-5472.Can-07-6538]
- Takasugi M, Okada R, Takahashi A, Virya Chen D, Watanabe S, et al. Small extracellular vesicles secreted from senescent cells promote cancer cell proliferation through EphA2. Nat Commun 2017;8:15728. [PMID: 28585531 DOI: 10.1038/ncomms15728]
- Pavlyukov MS, Yu H, Bastola S, Minata M, Shender VO, et al. Apoptotic cell-derived extracellular vesicles promote malignancy of glioblastoma via intercellular transfer of splicing factors. Cancer Cell 2018;34(1):119-35.e10. [PMID: 29937354 DOI: 10.1016/j.ccell.2018.05.012]
- Yang L, Wu XH, Wang D, Luo CL, Chen LX. Bladder cancer cell-derived exosomes inhibit tumor cell apoptosis and induce cell proliferation in vitro. Mol Med Rep 2013;8(4):1272-8. [PMID: 23969721 DOI: 10.3892/mmr.2013.1634]
- Branco H, Xavier CPR, Riganti C, Vasconcelos MH. Hypoxia as a critical player in extracellular vesicles-mediated intercellular communication between tumor cells and their surrounding microenvironment. Biochim Biophys Acta (BBA) – Rev Cancer 2025;1880:189244. [PMID: 39672279 DOI: 10.1016/j.bbcan.2024.189244]
- Chang WH, Cerione RA, Antonyak MA. Extracellular vesicles and their roles in cancer progression. Methods Mol Biol 2021;2174:143-70. [PMID: 32813249 DOI: 10.1007/978-1-0716-0759-6_10]
- Singh S, Chen CC, Kim S, Singh A, Singh G. Role of extracellular vesicle microRNAs and RNA binding proteins on glioblastoma dynamics and therapeutics development. Extracell Vesicle 2024;4:100049. [DOI: 10.1016/j.vesic.2024.100049]
- Apostolou S, Donega V. Embracing the heterogeneity of neural stem cells in the subventricular zone. Stem Cell Reports 2025;20(9):102452. [PMID: 40118056 DOI: 10.1016/j.stemcr.2025.102452]
- Zhao S, Umpierre AD, Wu LJ. Tuning neural circuits and behaviors by microglia in the adult brain. Trends Neurosci 2024;47(3):181-94. [PMID: 38245380 DOI: 10.1016/j.tins.2023.12.003]
- Xiao Y, Wang SK, Zhang Y, Rostami A, Kenkare A, et al. Role of extracellular vesicles in neurodegenerative diseases. Prog Neurobiol 2021;201:102022. [PMID: 33617919 DOI: 10.1016/j.pneurobio.2021.102022]
- Jezierzański M, Nafalska N, Stopyra M, Furgoł T, Miciak M, et al. Temozolomide (TMZ) in the treatment of glioblastoma multiforme—a literature review and clinical outcomes. Curr Oncol 2024;31(7):3994-4002. [PMID: 39057168 DOI: 10.3390/curroncol31070296]
- Ma S, Lu Y, Sui S, Yang JS, Fu BB, et al. Unraveling the triad of immunotherapy, tumor microenvironment, and skeletal muscle biomechanics in oncology. Front Immunol 2025;16:1572821. [PMID: 40242775 DOI: 10.3389/fimmu.2025.1572821]
- Chung WM, Molony RD, Lee YF. Non-stem bladder cancer cell-derived extracellular vesicles promote cancer stem cell survival in response to chemotherapy. Stem Cell Res Ther 2021;12(1):533. [PMID: 34627375 DOI: 10.1186/s13287-021-02600-6]
- Benmelouka AY, Munir M, Sayed A, Attia MS, Ali MM, et al. Neural stem cell-based therapies and glioblastoma management: current evidence and clinical challenges. Int J Mol Sci 2021;22(5):2258. [PMID: 33668356 DOI: 10.3390/ijms22052258]
- Zeng A, Wei Z, Rabinovsky R, Jun HJ, El Fatimy R, et al. Glioblastoma-derived extracellular vesicles facilitate transformation of astrocytes via reprogramming oncogenic metabolism. iScience 2020;23(8):101420. [PMID: 32795915 DOI: 10.1016/j.isci.2020.101420]
- Fattahi E, Kankam SB, Khoshnevisan A, Hashemi AP. Evaluating prognosis and survival in patients with glioblastoma in contact with subventricular zone: tumor location and its correlation with prognosis. Med J Armed Forces India 2024;80(Suppl 1):S21-8. [PMID: 39734827 DOI: 10.1016/j.mjafi.2022.06.012]
- Mendt M, Kamerkar S, Sugimoto H, McAndrews KM, Wu CC, et al. Generation and testing of clinical-grade exosomes for pancreatic cancer. JCI Insight 2018;3(8):e99263. [PMID: 29669940 DOI: 10.1172/jci.insight.99263]
- Luo X, McAndrews KM, Kalluri R. Natural and bioengineered extracellular vesicles in diagnosis, monitoring and treatment of cancer. ACS Nano 2025;19(6):5871-96. [PMID: 39869032 DOI: 10.1021/acsnano.4c11630]
- Hu M, Han Y, Zhang X, Tian S, Shang Z, et al. Extracellular vesicles for targeted drug delivery: advances in surface modification strategies and therapeutic applications. J Transl Med 2025;23(1):1028. [PMID: 41029680 DOI: 10.1186/s12967-025-07077-y]
- Patel N, LaMastro V, Giblin J, Avery E, Noor B, et al. Genetically engineering cells to produce therapeutically boosted extracellular vesicles for cardiovascular calcification. Biomaterials 2026;325:123552. [PMID: 40682948 DOI: 10.1016/j.biomaterials.2025.123552]
- Ye C, Ma Y, Shrestha R, Cai J, Liu Y, et al. Extracellular vesicle-mediated delivery of CRISPR machinery silences androgen receptor in castration-resistant prostate cancer cells. Mol Ther 2026;34(1):281-99. [PMID: 41017153 DOI: 10.1016/j.ymthe.2025.09.045]
- Park KC, Jaafari AM, Smith CA, Lobo AR, Errichelli L, et al. A Langendorff-heart discovery pipeline demonstrates cardiomyocyte targeting by extracellular vesicles functionalized with beta-blockers using click-chemistry. J MolCell Cardiol 2025;204:89-100. [PMID: 40414416 DOI: 10.1016/j.yjmcc.2025.05.007]
- Sato Y, Zhang W, Baba T, Chung UI, Teramura Y. Extracellular vesicle-liposome hybrids via membrane fusion using cell-penetrating peptide-conjugated lipids. Regen Ther 2024;26:533-40. [PMID: 39165408 DOI: 10.1016/j.reth.2024.07.006]
- Dave KM, Pinky PP, Manickam DS. Molecular engineering of extracellular vesicles for drug delivery: strategies, challenges, and perspectives. J Control Release 2025;386:114068. [PMID: 40721069 DOI: 10.1016/j.jconrel.2025.114068]
- Zhang T, Liu Z, Wei Y, Lu J, He Z, et al. Extracellular vesicles as natural nanocarriers: from in vitro engineering to in situ generation in cancer therapy. Chem Eng J 2025;510:161653. [DOI: 10.1016/j.cej.2025.161653]
- Ding L, Zhang T, Pan Y, Liu J, Ma T, et al. Extracellular vesicles in chronic wound therapy: engineering strategies and advanced delivery systems for enhanced regeneration. Mater Today Bio 2025;35:102298. [PMID: 41890412 DOI: 10.1016/j.mtbio.2025.102298]
- Mediratta K, Diab MD, Han P, Hu H, Wang L. Emerging strategies for cargo loading and engineering of extracellular vesicles for breast cancer treatment. Nanomaterials (Basel) 2025;15(18):1418. [PMID: 41003053 DOI: 10.3390/nano15181418]
- Liu Y, Xiang C, Dai Y, Li C, Okeke MN, et al. Engineered apoptotic vesicle mimetics with tunable “eat-me” signaling precisely regulate tumor-associated macrophages for potentiating cancer immunotherapy. Acta Pharm Sin B 2025. [DOI: 10.1016/j.apsb.2025.11.032]
- Zhang K, Fang J. Extracellular vesicles in drug delivery: from quality assurance to therapeutic application. Int J Nanomedicine 2026;21:579695. [DOI: 10.2147/IJN.S579695]
- Zhao C, Wang Z, Kim H, Kong H, Lee J, et al. Identification of tumor-specific surface proteins enables quantification of extracellular vesicle subtypes for early detection of pancreatic ductal adenocarcinoma. Adv Sci (Weinh) 2025;12(21):e2414982. [PMID: 40130819 DOI: 10.1002/advs.202414982]
- Omarini C, Catani V, Mastrolia I, Toss A, Banchelli F, et al. Extracellular vesicles-derived miR-21 as a biomarker for early diagnosis and tumor activity in breast cancer subtypes. Biomark Res 2025;13(1):14. [PMID: 39849610 DOI: 10.1186/s40364-025-00724-y]
- Ma S, Zhao N, Dong X, Wang Y, Song L, et al. Liquid biopsy-derived extracellular vesicle protein biomarkers for diagnosis and prognostic assessment of lung squamous cell carcinoma. Cancer Cell Int 2025;25(1):161. [PMID: 40275246 DOI: 10.1186/s12935-025-03792-0]
- Li LY, Liang SY, Cai MP, Ge JC, Tan HS, et al. Engineered extracellular vesicles as imaging biomarkers and therapeutic applications for urological diseases. Mater Today Bio 2025;32:101646. [PMID: 40160248 DOI: 10.1016/j.mtbio.2025.101646]
- Gao C, Zhao W, Feng R, Zhang L, Ge L, et al. Melanin nanoparticles-loaded lactobacillus fermentum exosomes for targeted and visualized treatment of ulcerative colitis. J Adv Res 2025;82:1049-67. [PMID: 40555281 DOI: 10.1016/j.jare.2025.06.068]
- Sun JX, Xia QD, Xu JZ, An Y, Ma SY, et al. A novel prostate cancer-specific fluorescent probe based on extracellular vesicles targeting STEAP1 applied in fluorescence guided surgery. J Control Release 2025;380:199-218. [PMID: 39894263 DOI: 10.1016/j.jconrel.2025.01.079]
- Yousefli Z, Rafiee N, Nourollahian T, Alipour A, Haghshenas Z, et al. Recent advances of circulating biomarkers with potential diagnostic, prognostic, and therapeutic value in pancreatic cancer: limitations of clinical application. Pathol Res Pract 2025;272:156045. [PMID: 40483840 DOI: 10.1016/j.prp.2025.156045]
- Jiang H, Kumarasamy RV, Pei J, Raju KRS, Kanniappan GV, et al. Integrating engineered nanomaterials with extracellular vesicles: advancing targeted drug delivery and biomedical applications. Front Nanotechnol 2025;6:1513683. [DOI: 10.3389/fnano.2024.1513683]
- Ma S, Dang D, Wang W, Wang Y, Liu L. Concentration optimization of combinatorial drugs using Markov chain-based models. BMC Bioinformatics 2021;22(1):451. [PMID: 34548014 DOI: 10.1186/s12859-021-04364-5]
- Ma S, Wu J, Liu Z, He R, Wang Y, et al. Quantitative characterization of cell physiological state based on dynamical cell mechanics for drug efficacy indication. J Pharm Anal 2023;13(4):388-402. [PMID: 37181289 DOI: 10.1016/j.jpha.2023.03.002]
- Kim SM, Yang Y, Oh SJ, Hong Y, Seo M, et al. Cancer-derived exosomes as a delivery platform of CRISPR/Cas9 confer cancer cell tropism-dependent targeting. J Control Release 2017;266:8-16. [PMID: 28916446 DOI: 10.1016/j.jconrel.2017.09.013]
- El Andaloussi S, Mäger I, Breakefield XO, Wood MJ. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discov 2013;12(5):347-57. [PMID: 23584393 DOI: 10.1038/nrd3978]
- Yang Z, Shi J, Xie J, Wang Y, Sun J, et al. Large-scale generation of functional mRNA-encapsulating exosomes via cellular nanoporation. Nat Biomed Eng 2020;4(1):69-83. [PMID: 31844155 DOI: 10.1038/s41551-019-0485-1]
- Xiao K, Lai Y, Yuan W, Li S, Liu X, et al. mRNA-based chimeric antigen receptor T cell therapy: basic principles, recent advances and future directions. Interdiscip Med 2024;2(1):e20230036. [DOI: 10.1002/INMD.20230036]
- Wang JH, Forterre AV, Zhao J, Frimannsson DO, Delcayre A, et al. Anti-HER2 scFv-directed extracellular vesicle-mediated mRNA-based gene delivery inhibits growth of HER2-positive human breast tumor xenografts by prodrug activation. Mol Cancer Ther 2018;17(5):1133-42. [PMID: 29483213 DOI: 10.1158/1535-7163.Mct-17-0827]
- Pi F, Binzel DW, Lee TJ, Li Z, Sun M, et al. Nanoparticle orientation to control RNA loading and ligand display on extracellular vesicles for cancer regression. Nat Nanotechnol 2018;13(1):82-9. [PMID: 29230043 DOI: 10.1038/s41565-017-0012-z]
- Zhang H, Wang J, Ren T, Huang Y, Liang X, et al. Bone marrow mesenchymal stem cell-derived exosomal miR-206 inhibits osteosarcoma progression by targeting TRA2B. Cancer Lett 2020;490:54-65. [PMID: 32682951 DOI: 10.1016/j.canlet.2020.07.008]
- Nie H, Xie X, Zhang D, Zhou Y, Li B, et al. Use of lung-specific exosomes for miRNA-126 delivery in non-small cell lung cancer. Nanoscale 2020;12(2):877-87. [PMID: 31833519 DOI: 10.1039/c9nr09011h]
- Katakowski M, Buller B, Zheng X, Lu Y, Rogers T, et al. Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett 2013;335(1):201-4. [PMID: 23419525 DOI: 10.1016/j.canlet.2013.02.019]
- Ohno S, Takanashi M, Sudo K, Ueda S, Ishikawa A, et al. Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol Ther 2013;21(1):185-91. [PMID: 23032975 DOI: 10.1038/mt.2012.180]
- Choi H, Choi Y, Yim HY, Mirzaaghasi A, Yoo JK, et al. Biodistribution of exosomes and engineering strategies for targeted delivery of therapeutic exosomes. Tissue Eng Regen Med 2021;18(4):499-511. [PMID: 34260047 DOI: 10.1007/s13770-021-00361-0]
- Agrawal AK, Aqil F, Jeyabalan J, Spencer WA, Beck J, et al. Milk-derived exosomes for oral delivery of paclitaxel. Nanomedicine 2017;13(5):1627-36. [PMID: 28300659 DOI: 10.1016/j.nano.2017.03.001]
- Kim MS, Haney MJ, Zhao Y, Mahajan V, Deygen I, et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine 2016;12(3):655-64. [PMID: 26586551 DOI: 10.1016/j.nano.2015.10.012]
- Yuan Z, Kolluri KK, Gowers KH, Janes SM. TRAIL delivery by MSC-derived extracellular vesicles is an effective anticancer therapy. J Extracell Vesicles 2017;6(1):1265291. [PMID: 28326166 DOI: 10.1080/20013078.2017.1265291]
- Li J, Li J, Peng Y, Du Y, Yang Z, et al. Dendritic cell derived exosomes loaded neoantigens for personalized cancer immunotherapies. J Control Release 2023;353:423-33. [PMID: 36470333 DOI: 10.1016/j.jconrel.2022.11.053]
- Tsering T, Nadeau A, Wu T, Dickinson K, Burnier JV. Extracellular vesicle-associated DNA: ten years since its discovery in human blood. Cell Death Dis 2024;15(9):668. [PMID: 39266560 DOI: 10.1038/s41419-024-07003-y]
- Ding N, Daci A, Krasniqi V, Butler R, Goddard A, et al. Engineered extracellular vesicles demonstrate altered endocytosis and biodistribution and have superior oral siRNA delivery efficiency compared to lipid nanoparticles. Int J Pharm X 2025;10:100428. [PMID: 41256914 DOI: 10.1016/j.ijpx.2025.100428]
- Chen Y, Liang S, Peng Y, Ma K, Yun K, et al. Nanomaterials-enabled mRNA delivery for cancer immunotherapy. Coord Chem Rev 2025;543:216945. [DOI: 10.1016/j.ccr.2025]
- Balaraman AK, Arockia Babu M, Afzal M, Sanghvi G, Rekha MM, et al. Exosome-based miRNA delivery: transforming cancer treatment with mesenchymal stem cells. Regen Ther 2025;28:558-72. [PMID: 40034540 DOI: 10.1016/j.reth.2025.01.019]
- Al-Khafaji MK, Rahbarizadeh F, Ahmadvand D. Oligoclonal anti-HER2 nanobody-targeted exosomes as a nano carrier for doxorubicin delivery to HER2-positive breast cancer in vitro and in vivo. Int J Biol Macromol 2025;333(Pt 2):148626. [PMID: 41213374 DOI: 10.1016/j.ijbiomac.2025.148626]
- Deng L, Zhang H, Zhang Y, Luo S, Du Z, et al. An exosome-mimicking membrane hybrid nanoplatform for targeted treatment toward Kras-mutant pancreatic carcinoma. Biomater Sci 2021;9(16):5599-611. [PMID: 34250995 DOI: 10.1039/d1bm00446h]
- Cheng Q, Shi X, Han M, Smbatyan G, Lenz HJ, et al. Reprogramming exosomes as nanoscale controllers of cellular immunity. J Am Chem Soc 2018;140(48):16413-7. [PMID: 30452238 DOI: 10.1021/jacs.8b10047]
- Shi X, Cheng Q, Hou T, Han M, Smbatyan G, et al. Genetically engineered cell-derived nanoparticles for targeted breast cancer immunotherapy. Mol Ther 2020;28(2):536-47. [PMID: 31843452 DOI: 10.1016/j.ymthe.2019.11.020]
- Wan C, Sun Y, Tian Y, Lu L, Dai X, et al. Irradiated tumor cell–derived microparticles mediate tumor eradication via cell killing and immune reprogramming. Sci Adv 2020;6(13):eaay9789. [PMID: 32232155 DOI: 10.1126/sciadv.aay9789]
- Lin W, Xu Y, Chen X, Liu J, Weng Y, et al. Radiation-induced small extracellular vesicles as “carriages” promote tumor antigen release and trigger antitumor immunity. Theranostics 2020;10(11):4871-84. [PMID: 32308755 DOI: 10.7150/thno.43539]
- Wang ZH, Peng WB, Zhang P, Yang XP, Zhou Q. Lactate in the tumour microenvironment: from immune modulation to therapy. EBioMedicine 2021;73:103627. [PMID: 34656878 DOI: 10.1016/j.ebiom.2021.103627]
- Nguyen Cao TG, Kang JH, Kim W, Lim J, Kang S, et al. Engineered extracellular vesicle-based sonotheranostics for dual stimuli-sensitive drug release and photoacoustic imaging-guided chemo-sonodynamic cancer therapy. Theranostics 2022;12(3):1247-66. [PMID: 35154485 DOI: 10.7150/thno.65516]
- Peng X, Peng Q, Wu M, Wang W, Gao Y, et al. A pH and temperature dual-responsive microgel-embedded, adhesive, and tough hydrogel for drug delivery and wound healing. ACS Appl Mater Interfaces 2023;15(15):19560-73. [PMID: 37036950 DOI: 10.1021/acsami.2c21255]
- Chen W, Li Y, Liu C, Kang Y, Qin D, et al. In situ engineering of tumor-associated macrophages via a nanodrug-delivering-drug (β-Elemene@Stanene) strategy for enhanced cancer chemo-immunotherapy. Angew Chem Int Ed Engl 2023;62(41):e202308413. [PMID: 37380606 DOI: 10.1002/anie.202308413]
- Cao Y, Wu T, Zhang K, Meng X, Dai W, et al. Engineered exosome-mediated near-infrared-II region V2C quantum dot delivery for nucleus-target low-temperature photothermal therapy. ACS Nano 2019;13(2):1499-510. [PMID: 30677286 DOI: 10.1021/acsnano.8b07224]
- Wen Z, Jiang L, Yu F, Xu X, Chen M, et al. scRNA-seq reveals NAMPT-mediated macrophage polarization shapes smooth muscle cell plasticity in pulmonary arterial hypertension. Interdiscip Med 2024;2(4):e20240016. [DOI: 10.1002/INMD.20240016]
- Naseri M, Bozorgmehr M, Zöller M, Ranaei Pirmardan E, Madjd Z. Tumor-derived exosomes: the next generation of promising cell-free vaccines in cancer immunotherapy. Oncoimmunology 2020;9(1):1779991. [PMID: 32934883 DOI: 10.1080/2162402x.2020.1779991]
- Gardiner C, Di Vizio D, Sahoo S, Théry C, Witwer KW, et al. Techniques used for the isolation and characterization of extracellular vesicles: results of a worldwide survey. J Extracell Vesicles 2016;5(1):32945. [PMID: 27802845 DOI: 10.3402/jev.v5.32945]
- Liang G, Zhu Y, Ali DJ, Tian T, Xu H, et al. Engineered exosomes for targeted co-delivery of miR-21 inhibitor and chemotherapeutics to reverse drug resistance in colon cancer. J Nanobiotechnology 2020;18(1):10. [PMID: 31918721 DOI: 10.1186/s12951-019-0563-2]
- Zhang X, Zhang H, Gu J, Zhang J, Shi H, et al. Engineered extracellular vesicles for cancer therapy. Adv Mater 2021;33(14):e2005709. [PMID: 33644908 DOI: 10.1002/adma.202005709]
- Gruenberg J, Maxfield FR. Membrane transport in the endocytic pathway. Curr Opin Cell Biol 1995;7(4):552-63. [PMID: 7495576 DOI: 10.1016/0955-0674(95)80013-1]
- Gudbergsson JM, Jønsson K, Simonsen JB, Johnsen KB. Systematic review of targeted extracellular vesicles for drug delivery – considerations on methodological and biological heterogeneity. J Control Release 2019;306:108-20. [PMID: 31175896 DOI: 10.1016/j.jconrel.2019.06.006]
- Gulati M, Grover M, Singh S, Singh M. Lipophilic drug derivatives in liposomes. Int J Pharm 1998;165(2):129-68. [DOI: 10.1016/S0378-5173(98)00006-4]
- Guo M, Wu F, Hu G, Chen L, Xu J, et al. Autologous tumor cell–derived microparticle-based targeted chemotherapy in lung cancer patients with malignant pleural effusion. Sci Transl Med 2019;11(474):eaat5690. [PMID: 30626714 DOI: 10.1126/scitranslmed.aat5690]
- Haney MJ, Klyachko NL, Zhao Y, Gupta R, Plotnikova EG, et al. Exosomes as drug delivery vehicles for Parkinson’s disease therapy. J Control Release 2015;207:18-30. [PMID: 25836593 DOI: 10.1016/j.jconrel.2015.03.033]
- Sayyed AA, Gondaliya P, Mali M, Pawar A, Bhat P, et al. MiR-155 inhibitor-laden exosomes reverse resistance to cisplatin in a 3D tumor spheroid and xenograft model of oral cancer. Mol Pharm 2021;18(8):3010-25. [PMID: 34176265 DOI: 10.1021/acs.molpharmaceut.1c00213]
- Haraszti RA, Miller R, Stoppato M, Sere YY, Coles A, et al. Exosomes produced from 3D cultures of MSCs by tangential flow filtration show higher yield and improved activity. Mol Ther 2018;26(12):2838-47. [PMID: 30341012 DOI: 10.1016/j.ymthe.2018.09.015]
- Xia P, Yuan H, Tian M, Zhong T, Hou R, et al. Surface-engineered extracellular vesicles with CDH17 nanobodies to efficiently deliver imaging probes and chemo-photothermal drugs for gastric cancer theragnostic. Adv Funct Mater 2022;33:2209393. [DOI: 10.1002/adfm.202209393]
- Wang D, Yao Y, He J, Zhong X, Li B, et al. Engineered cell-derived microparticles Bi2Se3/DOX@MPs for imaging guided synergistic photothermal/low-dose chemotherapy of cancer. Adv Sci 2019;7(3):1901293. [PMID: 32042550 DOI: 10.1002/advs.201901293]
- Yang K, Ren D, Wang Z, Dong Q, Xu M, et al. Apoptotic bodies encapsulating Ti2N nanosheets for synergistic chemo-photothermal therapy. Nanotechnology 2024;35(36):365703. [PMID: 38861968 DOI: 10.1088/1361-6528/ad5690]
- Zhang D, Qin X, Wu T, Qiao Q, Song Q, et al. Extracellular vesicles based self-grown gold nanopopcorn for combinatorial chemo-photothermal therapy. Biomaterials 2019;197:220-8. [PMID: 30669014 DOI: 10.1016/j.biomaterials.2019.01.024]
- Tian R, Wang Z, Niu R, Wang H, Guan W, et al. Tumor exosome mimicking nanoparticles for tumor combinatorial chemo-photothermal therapy. Front Bioeng Biotechnol 2020;8:1010. [PMID: 32984284 DOI: 10.3389/fbioe.2020.01010]
- Zhu Y, Li W, Lan F, Chen S, Chen X, et al. DNA nanotechnology in tumor liquid biopsy: enrichment and determination of circulating biomarkers. Interdiscip Med 2024;2(1):e20230043. [DOI: 10.1002/INMD.20230043]
- Zhu L, Dong D, Yu ZL, Zhao YF, Pang DW, et al. Folate-engineered microvesicles for enhanced target and synergistic therapy toward breast cancer. ACS Appl Mater Interfaces 2017;9(6):5100-8. [PMID: 28106372 DOI: 10.1021/acsami.6b14633]
- Wang C, Guan W, Peng J, Chen Y, Xu G, et al. Gene/paclitaxel co-delivering nanocarriers prepared by framework-induced self-assembly for the inhibition of highly drug-resistant tumors. Acta Biomater 2020;103:247-58. [PMID: 31846802 DOI: 10.1016/j.actbio.2019.12.015]
- Zhan Q, Yi K, Qi H, Li S, Li X, et al. Engineering blood exosomes for tumor-targeting efficient gene/chemo combination therapy. Theranostics 2020;10(17):7889-905. [PMID: 32685027 DOI: 10.7150/thno.45028]
- Baby HM, Zhang H, Selvadoss A, Pathrikar TV, Bajpayee AG. Rational design of extracellular vesicles for targeted drug delivery across physiological barriers. Nano Today 2026;66:102920. [DOI: 10.1016/j.nantod.2025.102920]
- Du X, Chen S, Meng T, Liu L, Li L, et al. Extracellular vesicles as precision therapeutic vectors: charting the future of cell-targeted therapies. Precis Med Eng 2025;2(2):100031. [DOI: 10.1016/j.preme.2025.100031]
- Li Q, Chen X, Xie J, Nie S. Engineered bacterial extracellular vesicles: developments, challenges, and opportunities. Engineering 2025;54:291-307. [DOI: 10.1016/j.eng.2025.06.042]
- Bader J, Brigger F, Leroux JC. Extracellular vesicles versus lipid nanoparticles for the delivery of nucleic acids. Adv Drug Deliv Rev 2024;215:115461. [PMID: 39490384 DOI: 10.1016/j.addr.2024.115461]
- Popowski KD, Moatti A, Scull G, Silkstone D, Lutz H, et al. Inhalable dry powder mRNA vaccines based on extracellular vesicles. Matter 2022;5(9):2960-74. [PMID: 35847197 DOI: 10.1016/j.matt.2022.06.012]
- Li Q, Xing H, Naeem A, Zhang K, Zheng A, et al. Extracellular vesicle-based mRNA therapeutics and vaccines. Exploration (Beijing) 2025;5(6):2240109. [PMID: 41476648 DOI: 10.1002/exp.20240109]
- Liang X, Gupta D, Xie J, Van Wonterghem E, Van Hoecke L, et al. Engineering of extracellular vesicles for efficient intracellular delivery of multimodal therapeutics including genome editors. Nat Commun 2025;16(1):4028. [PMID: 40301355 DOI: 10.1038/s41467-025-59377-y]
- Wang SM, Wang D, Shen YQ, Wang MH, Jia F, et al. Isolation and detection strategies for decoding the heterogeneity of extracellular vesicles. Chem Eng J 2025;507:160234. [DOI: 10.1016/j.cej.2025.160234]
- Picchio V, Pontecorvi V, Dhori X, Bordin A, Floris E, et al. The emerging role of artificial intelligence applied to exosome analysis: from cancer biology to other biomedical fields. Life Sci 2025;375:123752. [PMID: 40409585 DOI: 10.1016/j.lfs.2025.123752]
- Liu H, Geng Z, Su J. Engineered mammalian and bacterial extracellular vesicles as promising nanocarriers for targeted therapy. Extracell Vesicles Circ Nucl Acids 2022;3(2):63-86. [PMID: 39698442 DOI: 10.20517/evcna.2022.04]
- Chen H, Li Q. Recent advances in scalable exosome production: challenges and innovations. Chin J Plast Reconstr Surg 2025;7(3):149-63. [DOI: 10.1016/j.cjprs.2025.05.001]
- Zhong H, Mao Z, Li X, Deng A, Zhu Y, et al. A microfluidic device integrating magnetic stirring with tangential flow for high-efficiency isolation of small extracellular vesicles. Sens Actuators B Chem 2026;447(Pt 2):138898. [DOI: 10.1016/j.snb.2025.138898]
- Chen X, Liu X, Zhang C, Xia H, Qin X, et al. Isolation and detection of exosomes on microfluidic chips. Biomedical Instrumentation 2025;1(1):100007. [DOI: 10.1016/j.bmi.2025.100007]
- Ahn SH, Ryu SW, Choi H, You S, Park J, et al. Manufacturing therapeutic exosomes: from bench to industry. Mol Cells 2022;45(5):284-90. [PMID: 35534190 DOI: 10.14348/molcells.2022.2033]
- Mukerjee N, Bhattacharya A, Maitra S, Kaur M, Ganesan S, et al. Exosome isolation and characterization for advanced diagnostic and therapeutic applications. Mater Today Bio 2025;31:101613. [PMID: 40161926 DOI: 10.1016/j.mtbio.2025.101613]
- Jay SM. Addressing barriers to clinical translation of extracellular vesicle therapeutics. Mol Ther 2025;33(5):1879-80. [PMID: 40010335 DOI: 10.1016/j.ymthe.2025.02.020]
- Scharbert L, Strodel B. Innovative strategies for modeling peptide–protein interactions and rational peptide drug design. Curr Opin Struct Biol 2025;93:103083. [PMID: 40570517 DOI: 10.1016/j.sbi.2025.103083]
- Wen M, Wang J, Ou Z, Nie G, Chen Y, et al. Bacterial extracellular vesicles: a position paper by the microbial vesicles task force of the Chinese society for extracellular vesicles. Interdiscip Med 2023;1(3):e20230017. [DOI: 10.1002/INMD.20230017]
- Liu H, Zhang H, Han Y, Hu Y, Geng Z, et al. Bacterial extracellular vesicles-based therapeutic strategies for bone and soft tissue tumors therapy. Theranostics 2022;12(15):6576-94. [PMID: 36185613 DOI: 10.7150/thno.78034]
- Kang J, Wen J, Chen H, Zhu C, Bai Y. Progress of research on engineered extracellular vesicles from different sources for disease treatment. Histol Histopathol 2025;40(10):1501-18. [PMID: 40134377 DOI: 10.14670/HH-18-903]
- Fu P, Guo Y, Luo Y, Mak M, Zhang J, et al. Visualization of microRNA therapy in cancers delivered by small extracellular vesicles. J Nanobiotechnology 2023;21(1):457. [PMID: 38031152 DOI: 10.1186/s12951-023-02187-5]
- Zheng W, Zhu T, Tang L, Li Z, Jiang G, et al. Inhalable CAR-T cell-derived exosomes as paclitaxel carriers for treating lung cancer. J Transl Med 2023;21(1):383. [PMID: 37308954 DOI: 10.1186/s12967-023-04206-3]
- Liu X, Liu X, Luo X, Zhu M, Liu N, et al. Synergistic strategies for glioblastoma treatment: CRISPR-based multigene editing combined with immune checkpoint blockade. J Nanobiotechnology 2025;23(1):94. [PMID: 39920725 DOI: 10.1186/s12951-025-03112-8]
- Wu Y, Chen W, Deng J, Cao X, Yang Z, et al. Tumour-derived microparticles obtained through microwave irradiation induce immunogenic cell death in lung adenocarcinoma. Nat Nanotechnol 2025;20(8):1119-30. [PMID: 40389640 DOI: 10.1038/s41565-025-01922-3]
- Yang G, Li Z, Usman R, Liu Y, Li S, et al. From biogenesis to aptasensors: advancements in analysis for tumor-derived extracellular vesicles research. Theranostics 2024;14(10):4161-83. [PMID: 38994022 DOI: 10.7150/thno.95885]
- Kanada M, Kim BD, Hardy JW, Ronald JA, Bachmann MH, et al. Microvesicle-mediated delivery of minicircle DNA results in effective gene-directed enzyme prodrug cancer therapy. Mol Cancer Ther 2019;18(12):2331-42. [PMID: 31451563 DOI: 10.1158/1535-7163.Mct-19-0299]
- Mizrak A, Bolukbasi MF, Ozdener GB, Brenner GJ, Madlener S, et al. Genetically engineered microvesicles carrying suicide mRNA/protein inhibit schwannoma tumor growth. Mol Ther 2013;21(1):101-8. [PMID: 22910294 DOI: 10.1038/mt.2012.161]
- Zhao L, Gu C, Gan Y, Shao L, Chen H, et al. Exosome-mediated siRNA delivery to suppress postoperative breast cancer metastasis. J Control Release 2020;318:1-15. [PMID: 31830541 DOI: 10.1016/j.jconrel.2019.12.005]
- Kim G, Kim M, Lee Y, Byun JW, Hwang DW, et al. Systemic delivery of microRNA-21 antisense oligonucleotides to the brain using T7-peptide decorated exosomes. J Control Release 2020;317:273-81. [PMID: 31730913 DOI: 10.1016/j.jconrel.2019.11.009]
- Wang X, Zhang H, Bai M, Ning T, Ge S, et al. Exosomes serve as nanoparticles to deliver anti-miR-214 to reverse chemoresistance to cisplatin in gastric cancer. Mol Ther 2018;26(3):774-83. [PMID: 29456019 DOI: 10.1016/j.ymthe.2018.01.001]
- Erkan EP, Senfter D, Madlener S, Jungwirth G, Ströbel T, et al. Extracellular vesicle-mediated suicide mRNA/protein delivery inhibits glioblastoma tumor growth in vivo. Cancer Gene Ther 2017;24(1):38-44. [PMID: 27982017 DOI: 10.1038/cgt.2016.78]
- Cheng G, Li W, Ha L, Han X, Hao S, et al. Self-assembly of extracellular vesicle-like metal–organic framework nanoparticles for protection and intracellular delivery of biofunctional proteins. J Am Chem Soc 2018;140(23):7282-91. [PMID: 29809001 DOI: 10.1021/jacs.8b03584]
- Nie W, Wu G, Zhang J, Huang LL, Ding J, et al. Responsive exosome nano-bioconjugates for synergistic cancer therapy. Angew Chem Int Ed Engl 2020;59(5):2018-22. [PMID: 31746532 DOI: 10.1002/anie.201912524]
- Fan M, Liu H, Yan H, Che R, Jin Y, et al. A CAR T-inspiring platform based on antibody-engineered exosomes from antigen-feeding dendritic cells for precise solid tumor therapy. Biomaterials 2022;282:121424. [PMID: 35196606 DOI: 10.1016/j.biomaterials.2022.121424]
- Wang G, Hu W, Chen H, Shou X, Ye T, et al. Cocktail strategy based on NK cell-derived exosomes and their biomimetic nanoparticles for dual tumor therapy. Cancers (Basel) 2019;11(10):1560. [PMID: 31615145 DOI: 10.3390/cancers11101560]
- Yong T, Zhang X, Bie N, Zhang H, Zhang X, et al. Tumor exosome-based nanoparticles are efficient drug carriers for chemotherapy. Nat Commun 2019;10(1):3838. [PMID: 31444335 DOI: 10.1038/s41467-019-11718-4]
- Gong C, Tian J, Wang Z, Gao Y, Wu X, et al. Functional exosome-mediated co-delivery of doxorubicin and hydrophobically modified microRNA 159 for triple-negative breast cancer therapy. J Nanobiotechnology 2019;17(1):93. [PMID: 31481080 DOI: 10.1186/s12951-019-0526-7]
- Liu X, Zhang G, Yu T, Liu J, Chai X, et al. CL4-modified exosomes deliver lncRNA DARS-AS1 siRNA to suppress triple-negative breast cancer progression and attenuate doxorubicin resistance by inhibiting autophagy. Int J Biol Macromol 2023;250:126147. [PMID: 37544559 DOI: 10.1016/j.ijbiomac.2023.126147]
- Zhang H, Wu B, Wang Y, Du H, Fang L. Extracellular vesicles as mediators and potential targets in combating cancer drug resistance. Molecules 2025;30(3):498. [PMID: 39942602 DOI: 10.3390/molecules30030498]
- Qiu Y, Sun J, Qiu J, Chen G, Wang X, et al. Antitumor activity of cabazitaxel and MSC-TRAIL derived extracellular vesicles in drug-resistant oral squamous cell carcinoma. Cancer Manag Res 2020;12:10809-20. [PMID: 33149686 DOI: 10.2147/cmar.S277324]
- Zhu L, Wang C, Pang DW, Zhang ZL. Controlled release of therapeutic agents with near-infrared laser for synergistic photochemotherapy toward cervical cancer. Anal Chem 2019;91(10):6555-60. [PMID: 30994332 DOI: 10.1021/acs.analchem.8b05982]
- Kim S, Kang JH, Nguyen Cao TG, Kang SJ, Jeong K, et al. Extracellular vesicles with high dual drug loading for safe and efficient combination chemo-phototherapy. Biomater Sci 2022;10(11):2817-30. [PMID: 35384946 DOI: 10.1039/d1bm02005f]
- Chen M, Sun Y, Liu H. Cell membrane biomimetic nanomedicines for cancer phototherapy. Interdisciplinary Medicine 2023;1(2):e20220012. [DOI: 10.1002/INMD.20220012]
- Huang GT, Gronthos S, Shi S. Mesenchymal stem cells derived from dental tissues vs. those from other sources: their biology and role in regenerative medicine. J Dent Res 2009;88(9):792-806. [PMID: 19767575 DOI: 10.1177/0022034509340867]
- Jiang Z, Guan J, Qian J, Zhan C. Peptide ligand-mediated targeted drug delivery of nanomedicines. Biomater Sci 2019;7(2):461-71. [PMID: 30656305 DOI: 10.1039/c8bm01340c]
- Bellavia D, Raimondo S, Calabrese G, Forte S, Cristaldi M, et al. Interleukin 3- receptor targeted exosomes inhibit in vitro and in vivo Chronic Myelogenous Leukemia cell growth. Theranostics 2017;7(5):1333-45. [PMID: 28435469 DOI: 10.7150/thno.17092]
- Usman WM, Pham TC, Kwok YY, Vu LT, Ma V, et al. Efficient RNA drug delivery using red blood cell extracellular vesicles. Nat Commun 2018;9(1):2359. [PMID: 29907766 DOI: 10.1038/s41467-018-04791-8]
- Wang X, Mao K, Zhang X, Zhang Y, Yang YG, et al. Red blood cell derived nanocarrier drug delivery system: a promising strategy for tumor therapy. Interdiscip Med 2024;2(3):e20240014. [DOI: 10.1002/INMD.20240014]
- Yang Y, Wang X, Wang M, Xiang Z, Li X, et al. Dual genes manipulation enhanced chemotherapy potentiates antitumor immunity based on extracellular vesicle system for glioblastoma treatment. Chem Eng J 2024;500:156638. [DOI: 10.1016/j.cej.2024.156638]